





Сознание животных
Впервые опубликовано 23 декабря 1995 года; содержательно переработано 24 октября 2016 года.
Вопросы о сознании животных — в частности, у каких животных есть сознание и каким это сознание может быть, если оно вообще существует — и научные, и философские одновременно. Научные, поскольку ответы на них потребуют сбора информации при помощи научных методов. Кабинетные размышления, концептуальный анализ, логика, априорное построение теорий, трансцендентальные выводы или интроспекция — подобные методы не дадут нам информации о том, имеется ли в жизни утконоса, игуаны или кальмара (возьмем хотя бы их для примера) субъективный опыт. В определенный момент нам потребуется узнать что-то о животных.
Однако все эти вопросы являются также глубоко философскими и имеют эпистемологическое, метафизическое и феноменологическое измерения. Успех здесь, следовательно, неизбежно требует междисциплинарной работы философов, желающих опираться на эмпирические данные биологии животных, и ученых, восприимчивых к возникающим философским затруднениям.
Обоснование: выстраивание аналогий между животными и людьми
Существует множество причин, обуславливающих философский интерес к сознанию нечеловеческих животных (не относящихся к человеческому роду; далее — просто «животные»).
Во-первых, раз философия зачастую начинается с вопроса о месте человека в природе, один из способов, при помощи которого люди пытались определить свое место — сопоставление и противопоставление с теми явлениями природы, которые наиболее близки к ним, то есть с другими животными. На Западе, пожалуй, традиционный способ размышлять о сознании (и, возможно, по-прежнему наиболее очевидный для многих людей) заключается в том, чтобы рассматривать сознание как изначально врожденный дар человека, другие животные могут также иметь его — или не иметь, в зависимости от того, в достаточной ли степени они похожи на нас. Традиционная библейская космология говорит о том, что все животные возникли в результате намеренного божественного творения, и только человек был создан по подобию Бога, следовательно, он имеет особую, привилегированную роль в задуманном устройстве мироздания — к примеру, у него есть доступ к вечной жизни после смерти, в которой не будет избытка блох, муравьев и улиток. (Более глубокое рассмотрение того, как соотносится проблема наличия сознания у животных и христианская теология, см. в Lewis 2009: ch. 9.) Однако в рамках современной биологической картины мира люди, быть может, и уникальны в определенном (и, вероятно, весьма важном) отношении, но мы лишь один из многих видов животных — одна вершина одной ветви филогенетического дерева жизни, и не обладаем каким-то сугубо специальным статусом.
В рамках эволюционной перспективы сознание — это признак, которым обладают некоторые животные (по крайней мере у людей он есть). Среди возникающих вопросов можно перечислить следующие. Является ли сознание поздно сформировавшимся и узко распределенным признаком или же это довольно старый и широко распространенный признак? Сформировался ли этот признак только один раз или же несколько раз, причем независимым образом?
С этой точки зрения, вопрос «Обладают ли (нечеловеческие) животные сознанием?» звучит довольно странно, поскольку он, например, неявно объединяет летучих мышей и кроликов (как «нечеловеческих» животных) и противопоставляет их людям. В действительности же кролики гораздо ближе к людям, нежели к летучим мышам (Nishihara et al. 2006), так что подобная постановка вопроса содержит ложное допущение. Конечно, тезис, что человек — единственное животное, обладающее сознанием, согласуется с той перспективой, которую задает эволюционная теория. Это означало бы, что сознание возникло в результате недавнего эволюционного события, произошедшего после того, как наши генеалогические предки отделились от наших ближайших нечеловеческих родственников, шимпанзе и бонобо (обсуждение подобных гипотез см. в разд. 6). Однако такая точка зрения требует дополнительной аргументации в ее защиту; выбор этой позиции в качестве базовой, возможно и интуитивен для кого-то, но является произвольным.
Во-вторых, когда мы рассматриваем вопрос о том, являются ли животные существами сознательными или «бездумными автоматами», на карту поставлена мораль. (См. статью о моральном статусе животных.) Каждый год миллиарды животных забивают на мясо, используют в исследованиях, а также для других нужд человека. Более того, если считать, что опыт восприятия животных действительно соответствует опыту восприятия человека, то условия, в которых перед смертью находятся многие — пожалуй, даже большинство — животных, равносильны живодерству.
Когда рассматривается вопрос о сознании животных, дилемма о том, несем ли мы (как цивилизация) вину за огромную массу жестокости или являемся невиновными, также висит на волоске. Тем не менее некоторые философы утверждают, что сознание само по себе не играет роли в вопросах обращения с животными, а следовательно, либо (a) животные заслуживают рассмотрения с точки зрения морали, даже если они не имеют сознания, либо (б) животные не заслуживают рассмотрения с точки зрения морали, даже если они имеют сознание. (Другие разборы этических вопросов: Singer 1990 [1975]; Regan 1983; Rollin 1989; Varner 1998, 2012; Steiner 2008.)
В-третьих, хотя теории сознания зачастую разрабатываются без специального обращения к вопросам о сознании животных, их достоверность иногда оценивается на основе результатов их применения к сознанию животных (и, сходным образом, к сознанию детей). Это порождает вопросы об относительном эпистемологическом весе теоретических размышлений (напр., философских аргументов в пользу определенной теории сознания) в сравнении с суждениями или интуициями, касающимися конкретного случая, если нам надо решить, имеет ли данное существо сознание. Так, Серл (Searle 1998) утверждает, что наше интуитивное, соответствующее здравому смысле приписывание интенциональных и эмоциональных состояний собаке имеет больший эпистемологический вес, нежели философски обоснованные скептические доводы. Каррутерс (Carruthers 1989), напротив, заявляет, что его собственные аргументы в пользу того, что животные (даже собаки) не обладают сознанием, имеют достаточный вес, и что мы имеем моральное обязательство искоренять или игнорировать наше сопереживание в отношении таких существ. Должно ли наше интуитивное приписывание сознания животным (либо, напр., младенцам) ограничивать наши теории сознания или же теории должны идти наперекор этим интуициям?
В-четвертых, проблема определения того, обладают ли животные сознанием, сдвигает границы знания и научной методологии (за критический порог, по мнению некоторых). Так называемая когнитивная революция, которая имела место во второй половине XX века, повлекла за собой многочисленные инновационные эксперименты психологов-компаративистов и этологов, зондирующих когнитивные способности животных. Философские вопросы, возникающие при интерпретации экспериментов, направленных на исследование восприятия, обучения, категоризации, памяти, восприятия пространства, различения численности, коммуникации, языка, социального познания, теории психики (модели психического состояния, theory of mind), каузального мышления, а также метакогниций у животных, обсуждаются в статье, посвященной познавательной способности животных.
Несмотря на это исследование познавательной способности, вопрос о том, обладают ли животные сознанием самим по себе, остается для многих ученых дискуссионным и даже табуированным, в то время как другие ученые с самым разнообразным дисциплинарным опытом (напр., из нейробиологии, этологии, эволюционной биологии) разрабатывают новые способы рассмотрения данного вопроса (см. обзор: Boly et al. 2013).
Принятая в 2012 году Кембриджская Декларация о cознании демонстрирует, что многие ученые согласны с тем, что «совокупность свидетельств указывает на то, что люди не уникальны в обладании неврологическими субстратами, генерирующими сознание». Однако другие ученые, в том числе Мариан Стэмп Докинз, известная в науке о защите благополучия животных (Dawkins 1985, 1993), не готовы поддержать это заявление, и пишут следующее: «Тайна сознания остается. Пропасть в объяснении как всегда широка, и при всем желании мы не перешагнем ее» (Dawkins 2012: 171–172).
Многие философы и ученые или утверждали, или предполагали, что сознание заведомо индивидуально, а следовательно, опыт одного непостижим для других. Язык может позволить людям перешагнуть через эту предполагаемую пропасть посредством сообщения своего опыта другим, однако для других животных это по всей вероятности невозможно. Несмотря на расхождение мнений в философских и научных кругах, большинству людей здравый смысл говорит, что некоторые животные действительно имеют сознательный опыт. Если задать вопрос, почему знакомые нам животные, например наши домашние питомцы, обладают сознанием, большинство людей укажет на сходство в поведении этих животных и человека: так, нам кажется, что животные наглядно выражают удовольствие и неудовольствие, а также разнообразные эмоции, что их поведение мотивировано поиском пищи, комфорта, социальных контактов и т.д., что они осознают свое окружение и способны учиться на опыте. Аргументы сходства, используемые в пользу наличия сознания у животных, коренятся, таким образом, в наблюдениях, основанных на здравом смысле. Однако научные исследования поведения и компаративистские исследования анатомии мозга и психологии, а также соображения эволюционной преемственности между видами также могут подкреплять эти аргументы.
Даже при наличии структурных различий два мозга могут быть схожи с точки зрения нейродинамики, что позволяет сделать выводы о наличии сознания у животных (Seth et al. 2005).
Не только общие доводы о связи между сознанием, нейронной активностью и поведением, но и значительное количество научных исследований, направленных на понимание конкретных сознательных состояний, прибегают к животным как к заместителям человека. Реакции многих животных (особенно других млекопитающих) на телесные события, которые человек квалифицировал бы как болезненные, автоматически и легко признаются большинством людей реакциями на боль. Издавание высоких звуков, реакции страха, лечение ран, выученное избегание — это примеры реакций на болевые раздражители, все они входят в состав общего наследия млекопитающих, и сходные реакции также наблюдаются у организмов, относящихся к самым различным таксономическим группам (см. разд. 7.1 ниже).
Большая часть исследований, имеющих непосредственное отношение к лечению человеческой боли, включая исследования эффективности обезболивающих и анестетиков, проводится на крысах и других животных.
Валидность таких исследований основывается на схожести вовлеченных механизмов [1], и отрицание того, что раненые крысы, к которым применяются, например, опиаты, испытывают боль, кажется многим необоснованным [2]. Сходным образом большая часть фундаментальных исследований, имеющих непосредственное отношение к пониманию человеческого зрительного сознания, проводится на обезьянах, имеющих очень похожую зрительную систему. Обезьяны с поврежденной первичной зрительной корой демонстрируют нарушения, аналогичные нарушениям у пациентов со слепозрением (Stoerig & Cowey 1997), из чего можно сделать вывод, что зрительное сознание обезьян, не подвергшихся воздействию в ходе эксперимента, сходно со зрительным сознанием интактных людей.
Научные иллюстрации того, что представители других видов, даже других типов, подвержены тем же зрительным иллюзиям, что и мы (напр., Fujita et al. 1991), позволяют предположить сходство их зрительного опыта.
Часто говорят о том, что использование животных для моделирования нейропсихиатрических расстройств предполагает сходство эмоциональных и других сознательных состояний, и дальнейшее совершенствование такого моделирования может усилить аргумент в пользу приписывания подобных состояний животным. Любопытный разворот диспозиции моделирования можно найти в работе Темпл Грандин, профессора животноводства в университете штата Колорадо: в качестве основы ее понимания природы животного опыта выступает ее собственный опыт так называемой высокофункциональной аутистки (Grandin 1995, 2004).
Такие аргументы сходства, конечно, изначально ограничены, поскольку всегда дают возможность критикам эксплуатировать те или иные различия между животными и людьми и возражать, что наличие сходства не означает наличия у животных такой же чувствительности, как та, что имеется у человека. Даже в тех случаях, когда аргументы подкрепляются эволюционными соображениями о преемственности между видами, они остаются уязвимыми, поскольку сам по себе факт наличия у человека определенного признака не означает, что наши ближайшие родственники также должны иметь этот признак. Утверждение, что только человек способен научиться играть в шахматы, никак не противоречит эволюционной преемственности. То же верно и в отношении сознания. Повинелли и Джамброун (Povinelli & Giambrone 2000) также утверждают, что аргумент, основанный на аналогии, проваливается, поскольку поверхностное наблюдение за весьма схожим поведением даже близкородственных видов не гарантирует того, что когнитивные принципы, лежащие в его основе, являются одинаковыми. Именно это, с точки зрения Повинелли, демонстрируется в его исследовании того, как шимпанзе используют подсказки для отслеживания зрительного внимания (Povinelli 1996).
Возможно, сочетание поведенческих, физиологических и морфологических сходств с эволюционной теорией эквивалентно более сильному общему доводу [3]. Однако убедительный аргумент, вероятно, также требует обоснования в виде хорошо разработанной теории, описывающей структуру и функционирование сознания как когнитивного процесса — этот путь избрали многие недавние участники споров о сознании животных (см. разд. 6).
Концепции сознания
Термин «сознание» печально известен своей неоднозначностью, и определить его довольно трудно. Произрастая из народной, или обыденной, психологии (folk psychology), термин «сознание» имеет многочисленные употребления, которые могут и не отображаться в рамках единой когерентной концепции (Wilkes 1984). Тем не менее среди разных понятий сознания было выявлено несколько полезных отличий, которые позволяют немного прояснить часть важных вопросов о сознании животных.
Существует два обыденных понимания сознания, неоспоримые в случае их применения к животным: понимание сознания, которое имеет место, если существо бодрствует, а не находится во сне [4] или в коме, а также понимание сознания, которое вытекает из фундаментальной способности организмов воспринимать и благодаря этому реагировать на определенные характеристики их среды, что позволяет говорить о том, что животное является сознательным или осведомлено об этих характеристиках.
Сознание в обоих указанных смыслах может быть выявлено у организмов, представляющих самые разнообразные таксоны (см., напр., Mather 2008).
Третье, более специальное понимание сознания — так называемое сознание доступа — было предложено Блоком (Block 1995), чтобы зафиксировать тот смысл, в котором ментальные репрезентации могут использоваться при рациональном контроле действий или речи. Такое «диспозициональное» описание сознания доступа — мысль, что репрезентативное содержание доступно для использования другими системами, — было усовершенствовано Блоком (Block 2005), чтобы включить еще один возникший аспект — что содержание «транслируется» в «глобальном рабочем пространстве» (Baars 1997), которое затем становится доступным задач, требующих более высокой когнитивной обработки, таких как категоризация, рассуждение, планирование и произвольное направление внимания. Блок уверен, что многие животные обладают сознанием доступа (речь не является необходимым требованием). Действительно, некоторые свидетельства из области неврологии, которые цитирует Блок (Block 2005) для подкрепления тезиса о глобальном рабочем пространстве, получены в ходе экспериментов с обезьянами. Однако очевидно, что такой автор, как Декарт, который, как мы увидим, отрицал наличие речи, языка и рациональности у животных, отрицал бы и наличие у них сознания доступа. Те, кто вслед за Девидсоном (Davidson 1975) отрицают наличие интенциональных состояний у животных, скорее всего согласятся с этим.
Остается еще два понимания сознания, применительно к животным они вызывают еще больше споров: феноменальное сознания и самосознание.
Феноменальное сознание ссылается на квалитативные, субъективные, опытные или феноменологические аспекты сознательного опыта, которые иногда отождествляются с квалиа. (В данной статье мы также используем термин «чувствительность» в отношении феноменального сознания).
Нагель (2003) оспаривает нашу способность знать, вообразить или описать в научных (объективных) терминах то, каково быть летучей мышью, однако он считает, что есть нечто, на что это похоже.
Для многих авторов формулировка «каково быть», которую использует Нагель для феноменального сознания, служит опорной точкой, указывающей, что стоит на кону в спорах о сознании животных. В исследовании того, обладает ли группа животных сознанием, принципиальный вопрос — действительно ли «быть [этими животными] на что-то похоже», то есть существует ли для них субъективный опыт жизни или существования, собственная перспектива, которая есть у индивидов в отношении присущих им перцептивных, когнитивных и эмоциональных процессов.
Хотя некоторые авторы (включая самого Нагеля) утверждают, что именно субъективность феноменального сознания делает чрезвычайно сложным или практически невозможным его научное исследование, особенно у других видов, другие продолжают развивать структурные и/или функциональные теории сознания и используют их, отстаивая ту или иную гипотезу о распределении сознания среди животных. Подобные теории будут обсуждаться ниже, в разд. 5 и 6.
Термин самосознание указывает на то, что субъект осведомлен о себе, но он также печально известен своей неоднозначностью — субъект может быть осведомлен о себе в принципиально разных смыслах (см., напр., статью о феноменологических теориях самосознания).
К ним относятся: быть осведомленным о собственном теле как физическом объекте или как в посреднике своего восприятия или действия (т.е. телесная осведомленность о себе); быть осведомленным о своих ментальных состояниях (т.е. ментальная или опытная осведомленность о себе); быть осведомленным о том, что тебя воспринимают другие (т.е. социальная осведомленность о себе); быть осведомленным о себе как об устоявшемся герое собственного нарратива и нарратива других (т.е. нарративная осведомленность о себе). Это далеко не исчерпывающий список, и в случае каждого указанного понятия необходимо дальнейшее преодоление многозначности. Следовательно, хотя по многим теориям самосознание тесно связано с феноменальным сознанием, теории, выдвигающие аргументы в пользу их связи, могут существенно варьироваться с точки зрения их значения и следствий для вопроса о том, какие животные могли бы обладать сознанием.
Оставшаяся часть статьи рассматривает главным образом приписывание сознания в его феноменальном смысле животным, хотя в ней будут также обсуждаться вопросы сознания доступа, самосознания и теории психики применительно к животных, особенно когда эти вопросы имеют теоретическое отношение к феноменальному сознанию — как, например, в случае аргумента Каррутерса (Carruthers 1998a,b, 2000), согласно которому для феноменального сознания необходим особый вид ментальной саморепрезентации.
История вопроса
Как нам представляется сквозь фильтр «нововременной» философии Декарта, вопросы о наличии сознания у животных в западной традиции восходят к античным спорам о природе человека. Было бы анахронизмом толковать идеи, касающиеся сознания, представленные в античных источниках, с точки зрения наших дней.
Аристотель утверждал, что только у человека имеется разумная душа, а душа как движение присуща всем животным — как тем, которые относятся к человеческому роду, так и тем, которые к нему не относятся — и наделяет животных инстинктами, предназначенными для их успешного размножения и выживания. Сорабджи утверждает, что отрицание разума у животных породило кризис греческой мысли и потребовало «тотального переосмысления» природы ментальных способностей, а также пересмотра способа размышления о «человеке и его месте в природе, превосходящем животных» (Sorabji 1993: 7).
Дискуссии о том, что есть мышление и демонстрируют ли его животные, остаются с нами и 25 столетий спустя, о чем свидетельствует издание Rational Animals? (Hurley & Nudds 2006). Великая цепь бытия, выведенная из раннехристианской трактовки аристотелевских звеньев природы (Лавджой 2001), — еще один пример влияния аристотелизма на споры о психике животных.
Спустя два тысячелетия после Аристотеля механистическая философия Декарта для объяснения поведения животных ввела представление о рефлексе. Хотя его концепция рассматривала животное как машину, движимую рефлексами и не обладающую интеллектуальными способностями, важно признать, что его механистическое объяснение было абсолютно адекватным объяснением ощущений и восприятий — тех аспектов поведения животных, которые сегодня часто ассоциируются с сознанием. Граница была проведена только в отношении рационального мышления и понимания. Если принять во внимание и аристетелевское различение инстинкта и мышления, и картезианское различение механического рефлекса и рациональной мысли, то возникает соблазн установить соответствие между ними. Несмотря на это, уподобление данных оппозиций может оказаться ошибочным. Во-первых, целый ряд авторов прежде и после Дарвина полагали, что инстинктивные и рефлекторные действия могут сопровождаться сознательным опытом. Во-вторых, зависимость феноменального сознания от рационального и саморефлексивного мышления — довольно сильный и спорный тезис (хотя в настоящий момент у него есть защитники, что будет обсуждаться ниже).
Хотя тщательные наблюдения и эксперименты над природным миром берут истоки еще в античности, изучение поведения животных еще долгое времени после научной революции оставалось в основном бессистемным. Конечно, животных широко использовали в поисках ответов на анатомические, физиологические и эмбрионологические вопросы. Вивисекции проводились такими светилами античности, как Гален, а в начале Нового времени эта практика была возрождена (Bertoloni Meli 2012). Сам Декарт практиковал вивисекцию и выступал в ее защиту (письмо Декарта Племпию от 15 февраля 1638 года), в переписке он упоминал, что механистическое понимание животных освобождает людей от какой бы то ни было вины за убийство животных и употребление их в пищу. Последовавшие за ним механицисты (напр., Мальбранш) использовали тезис Декарта об отсутствии разума и души у животных для того, чтобы обосновать свое убеждение, что животные неспособны страдать или испытывать эмоции, а также не заслуживают отношения с точки зрения морали; они также оправдывали вивисекцию и другие виды жестокого обращения с животными (см. Olson 1990: 39–40).
Некоторые проблески экспериментальных подходов к поведению животных можно увидеть в конце XVIII века (напр., Barrington 1773; White 1789). Вскоре после этого, с 1804 года и вплоть до своей смерти в 1838 году Фредерик Кювье начал проводить исследования формирования сексуального и социального поведения у млекопитающих, обитающих в неволе. В середине XIX века Альфред Рассел Уоллес (Wallace 1867) уже открыто выступал за экспериментальный подход в отношении поведения животных, а эксперименты Дугласа Сполдинга (Spalding 1872), касающиеся инстинктивного пищевого поведения цыплят, стали культовыми. Тем не менее появившиеся экспериментальные подходы мало что могли сказать о сознании самом по себе, хотя можно считать, что работы Сполдинга внесли вклад в дискуссии об инстинкте и разуме.
Противопоставляя таким же образом инстинкт и разум, в своей работе
«Происхождение человека» Дарвин писал:
Как в «Происхождении видов», так и в «Происхождении человека» он уделяет существенное внимание поведению животных, очевидно стремясь продемонстрировать ментальную преемственность между видами. Доказывая свою правоту, Дарвин всецело полагался на единичные наблюдения (anecdotes) своих собеседников, излагаемые ими в переписке, — этот проект, дурно воспринятый в дальнейшем, продолжил после смерти Дарвина его протеже Джордж Роменс (Romanes 1882). При этом Дарвин также ставил опыты и был заядлым наблюдателем. В своей последней работе он описывает эксперименты, касающиеся пластичности поведения дождевого червя при обращении с листьями, которые проводились с тем, чтобы показать изрядную разумность (Darwin 1881; см. также Crist 2002).
Представление о пластичности поведения (behavioral flexibility) занимает центральное место в дискуссиях о психике и сознании животных. Картезианское понимание животного как автомата превращает феноменальное сознание не более чем в излишек. Философское развитие этой связки проследил Т. Г. Гексли (Huxley 1874). Гексли описал ряд опытов над лягушкой, в которых демонстрируется сходное рефлекторное поведение даже в том случае, когда спинной мозг лягушки был рассечен или у нее были удалены большие участки головного мозга. Он утверждал, что без мозга лягушка не могла бы находиться в сознании, однако поскольку она по-прежнему могла совершать то же, что и раньше, нет необходимости предполагать наличие сознания даже в случае наличия всего мозга, и продолжал настаивать на том, что сознание является излишком.
Тем не менее для тех (включая Гексли), кто быстро убедился в состоятельности дарвиновской теории эволюции, на первый план вышли понимание и защита ментальной преемственности между человеком и животными. Уильям Джеймс в «Принципах психологии» (James 1890) продвигал представление о различной интенсивности сознательного опыта среди животного царства. Эту идею воспроизвел ведущий британский психолог своего времени Конви Ллойд Морган в учебнике «Введение в сравнительную психологию» (An Introduction to Comparative Psychology, Lloyd Morgan 1894). Морган был настроен крайне скептически и критически в отношении «анекдотического подхода», которому благоволили Дарвин и Романес, однако он соглашается если не с дарвиновской методологией, то с его точкой зрения по вопросу ментальной преемственности. Стремясь устранить методологический дефицит, он ввел свой метод «двойной индукции» для понимания ментальных состояний животных (Lloyd Morgan 1894). Двойная индукция состояла из индуктивных выводов, основанных на наблюдении за поведением животных, в сочетании с интроспективным знанием нашей собственной психики. В то же время, чтобы противодействовать антропоморфному искажению в методе двойной индукции, Ллойд Морган ввел принцип, известный теперь как канон Моргана:
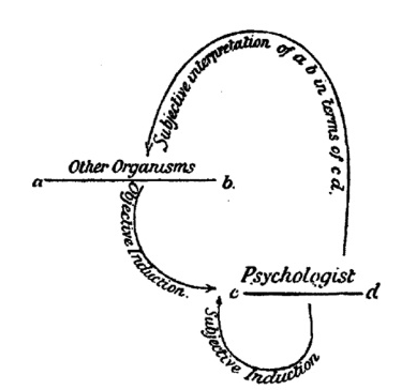
Иллюстрация метода двойной индукции Ллойда Моргана из его учебника 1894 года.
[cd — психолог, ab — другие организмы; сверху — субъективная интерпретация ab с точки зрения cd; снизу слева — объективная индукция, справа — субъективная индукция]
Несмотря на то, что двойной индуктивный метод сейчас представляет в основном лишь исторический интерес, канон Моргана жив и по сей день. Вопросы о том, что именно означает канон и как его обосновать, составляют предмет активных исторических и философских исследований (напр., Burghardt 1985; Sober 1998, 2005, 2012; Radick 2000; Thomas 2001, Fitzpatrick 2008). К ним также относятся вопросы о том, что Ллойд Морган имеет в виду под характеристиками «высшая» и «более низкая», в какой степени принцип может или должен быть обоснован эволюционными соображениями, а также сводится ли канон к принципу экономии (разновидности бритвы Оккама) или же к некоторым общим принципам эмпирического обоснования. Несмотря на имеющуюся неопределенность в отношении того, что на самом деле означает канон Моргана, будучи истолкованным (неверно — с точки зрения Thomas 2001) в качестве сильной версии принципа экономии, он служил центральной риторической фигурой для психологов бихевиористского толка, которые стремились исключить из сравнительной психологии любой намек на картезианский дуализм.
В американской психологии начала XX века господствовал бихевиоризм начиная с экспериментов Торндайка (Thorndike 1911) с животными, которые методом проб и ошибок обучались выбираться из сконструированных им «проблемных ящиков». Однако даже знаменитый «закон эффекта» Торндайка ссылается на «удовлетворение или дискомфорт» животного (Thorndike 1911: 244). Американская психология стала в первую очередь наукой о поведении, а не наукой о психике и поведении, как это было бы сформулировано в словаре, благодаря радикальному антиментализму Джона Уотсона (Watson 1928) и Б. Ф. Скиннера (Skinner 1953) — оба решительно отвергали любую попытку объяснить поведение животных в терминах ненаблюдаемых ментальных состояний.
В то же время в Европе все развивалось иначе, там преобладали этологические подходы к поведению животных. Этология — это отчасти естественная история с упором на полевые исследования, отчасти экспериментальная наука, которая имеет дело с животными, содержащимися в неволе. В ней нашли отражение стили двух знаковых для дисциплины фигур — Конрада Лоренца и Николаса Тинбергена (см. Burkhardt 2005). Изначально в своей работе Лоренц фокусировался на «врожденном» поведении. С его точки зрения, именно исследование врожденных способов поведения у родственных видов ставит изучение поведения животных в один ряд с другими направлениями эволюционной биологии. Он демонстрировал возможность вывести филогенетические отношения между видами посредством сравнения наборов их инстинктивных поведенческих схем (Lorenz 1971a). Следуя в этом направлении, Лоренц и Тинберген открыто стремились дистанцировать этологию от целеполагающей, менталистской психологии животных Биренса де Хана, а также от американской компаративисткой психологии, которой, с их точки зрения, недоставало внимания к биологии (см. Brigandt 2005). Подобно Ллойду Моргану, этологи отрицали анекдотический метод Роменса, но также критиковали и субъективистский подход самого Ллойда Моргана.
В 1970-х годах Дональд Гриффин, получивший известность благодаря точным физическим измерениям, доказывающим, что летучие мыши используют эхолокацию, произвел настоящий фурор своим призывом вернуться к вопросу о наличии психики у животных и особенно к вопросу о наличии у них сознания.
Гриффин ввел термин «когнитивная этология», чтобы описать такую исследовательскую программу, которая основывается на натуралистических наблюдениях поведения животных и на попытке понять психику животных в контексте эволюции.
Резкая критика в отношении Гриффина (Griffin 1978) появилась как со стороны психологов, так и со стороны классически вышколенных этологов.
Гриффин делал акцент на пластичности и переменчивости как на главных свидетельствах в пользу сознания, которое он определял как «субъективное состояние чувствования или рамышления об объектах и событиях» (Griffin & Speck 2004: 6). Позиция Гриффина также имеет существенное сходство с Ллойдом Морганом, поскольку он рассматривает субъективность, по крайней мере в простых формах, как феномен, широко распространенный в царстве животных. Бургхардт сообщает, что в ходе Далемской конференции, которую Гриффин созвал на ранней стадии формирования дисциплины, возникла «существенная неловкость с субъективизмом» (Burghardt 1985: 907).
Субъективистская позиция Гриффина и предположение, что даже такие насекомые, как пчелы, обладают сознанием, казались многим ученым досадным возвращением к антропоморфическому додумыванию на основе анекдотических свидетельств, виденному ранее у Дарвина и Роменса. Возможно, эта критика частично была несправедлива, поскольку Гриффин не повторяет истории в духе «друга фермера», которые собирал Роменс, но основывает свои интерпретации на результатах, приводимых в более взыскательной научной литературе, накопленной более чем за столетие после Дарвина (напр., Giurfa et al. 2001).
Тем не менее обвинения в додумывании результатов здесь избежать довольно сложно. Также важно отметить роль нейрологических свидетельств в его аргументах, когда он делает вывод, что активный поиск нейронных коррелятов сознания не продемонстрировал «никакой структуры или процесса, необходимых для сознаниях, которые обнаруживались бы исключительно в мозгу человека» (Griffin & Speck 2004). Эта точка зрения широко распространена среди нейроученых, однако не является общепризнанной.
От методологии Гриффина, основывающейся на поведении животных при изучении их сознания, также отказались как от антропоморфной (см. исследование Bekoff & Allen 1997). Но, возможно, опасность антропоморфизма переоценивается (Fisher 1990), и многие критики сами полагаются на такие утверждения, в пользу которых имеются лишь скудные научные данные (напр., Kennedy 1992, который утверждает, что «грех» антропоморфизма может быть генетически запрограммирован в человеке).
Одновременно с этим другие ученые, вне зависимости от того, одобряли они программу Гриффина или нет, стремились расширить эволюционные исследования сознания животных и включить в них нейронауки, а также широкий спектр функциональных соображений (напр., Ârhem et al. 2002, также см. разд. 6 ниже). Каковы бы ни были недостатки конкретных предложений Гриффина, он сыграл решающую роль в возобновлении развернутых дискуссий о сознании в науке о поведении и познании животных и тем самым открыл путь для современных исследований распределения сознания и его эволюционных истоков.
Эпистемологические и метафизические аспекты
Проблематика наличия сознания у нечеловеческих животных для изучающих сознание философов представляет в первую очередь эпистемологический интерес. Здесь имеют места два главных вопроса:
1. Можем ли мы знать, какие животные помимо человека обладают сознанием? (вопрос распределения) [5]
2. Можем ли мы знать, что представляет собой опыт животного, если он вообще существует? (феноменологический вопрос)
Томас Нагель в культовом эссе «Каково быть летучей мышью?» (Нагель 2003) просто предполагает, что существует нечто соответствующее тому, каково быть летучей мышью, и сосредотачивается на проблеме знания того, каково это — не поддающейся, с его точки зрения, научному решению. Уверенность Нагеля в наличии сознательного опыта у летучих мышей будет рассматриваться в целом как точка зрения, основанная на здравом смысле, которую многие ученые по-прежнему чаще всего воспринимают как нечто само собой разумеющееся, что демонстрирует предыдущий раздел. Однако, как мы увидим, ее можно оспорить, и есть те, кто утверждает, что вопрос распределения не поддается научному разрешению в той же степени, что и феноменологический вопрос.
Проблема других сознаний
Два указанных выше вопроса можно рассматривать как особые случаи — или, напротив, как обобщенные версии — скептической «проблемы других сознаний». Как можно узнать, что у других есть ментальные состояния, подобные нашим? Несмотря на отсутствие общепризнанного решения проблемы других сознаний, психологи спокойно игнорируют ее, как, по факту, и большинство людей, стремящихся на практике считать само собой разумеющимся то, что другие имеют ментальные состояния, подобные их собственным.
Несмотря на попытки обучить представителей других видов языкам, аналогичным человеческим, ни одна из них не достигла того уровня, который бы продемонстрировал способность вести разговор, что позволило бы непосредственно решить проблему (см. обзор Anderson 2004).
Кроме того, за исключением некоторых исследований, связанных с языком у попугаев и дельфинов, подобные подходы, как правило, ограничиваются теми животными, которые больше всего похожи на нас самих, в частности высшими приматами. Однако существует большая заинтересованность в возможных формах сознания, присущих гораздо более широкому спектру видов в сравнении с тем, что подходит для подобных исследований.
В более общем смысле проблема других сознаний становится более острой в случае нечеловеческих животных, поскольку сходство поведения и тела человека с поведением и телом других животных (что составляет основу решений проблемы «по аналогии») становится менее буквальным. Кроме того, перцептивный доступ к другому сознанию, который, как утверждают некоторые, сглаживает проблему, в отношении умов других животных выражен гораздо меньше. (Sober 2000 обсуждает проблему других сознаний в рамках эволюционного подхода, а Farah 2008 описывает перспективу нейронаук).
Эпистемический статус интуиций и восприятия ментальных состояний
Многим людям кажется очевидным, что знакомые им животные, такие как собаки и кошки, имеют сознательную ментальную жизнь, которая включает состояния восприятия, побуждения и наслаждения, базовые эмоции и социальные привязанности. Дэвид Юм, известный благодаря отстаиванию скептицизма, писал, что «ни одна истина не кажется мне более очевидной, чем там, что не только люди, но и животные обладают способностью мышления и рассудка» (Юм 1996: 228). Юм не приводит скрупулезных философских или эмпирических доводов в пользу тезиса — он полагал, что тот и так явственно вытекает из наблюдений. Серл формулирует его следующим образом:
Я делаю вывод о том, что моя собака обладает сознанием, не с бόльшим основанием, нежели, войдя в эту комнату, делаю вывод о том, что присутствующие люди обладают сознанием. Я просто реагирую на них так, как полагается в случае сознательных существ. Я просто отношусь к ним как к сознательным существам, и все. (Searle 1998)
Сторонники теорий, отрицающих наличие сознания у таких животных, должны отвергать приписывание эпистемического веса подобным интуициям. Например, Дэниел Деннет (чья позиция гласит, что сознание присуще исключительно человеку) считает, что интуитивное приписывание психических состояний «ненадежно», и подчеркивает:
Каррутерс признает, что интуитивное приписывание сознания животным широко распространено и идет рука об руку с сострадательным отношением к животным (напр., желанием предотвратить страдания). Однако он утверждает, что подобное сострадательное отношение некорректно, и наш моральный долг — ликвидировать или по крайней мере обойти его:
Необходимо отметить, что хотя Каррутерс продолжает утверждать, что только люди обладают сознанием, в последнее время он скорректировал свои этические взгляды, считая, что животные, несмотря на отсутствие сознания, возможно, в некоторой степени заслуживают отношения с точки зрения морали (Carruthers 1999).
Принципиальный момент здесь составляет мера надежности дотеоретических интуиций, касающихся психики животных. Назовем точку зрения, в соответствии с которой существует прямое восприятие (по крайней мере некоторых) ментальных состояний других, перцептуализмом. Он гласит, что по крайней мере иногда, когда мы наблюдаем ментальное состояние другого (напр., он радуется или испытывает боль), его ментальное состояние входит в содержание нашего восприятия — мы воспринимаем ментальные состояния других людей. Противоположную точку зрения, в соответствии с которой мы воспринимаем «исключительно поведение» и должны выводить или обосновывать наличие ментальных состояний, назовем инференциализмом.
Альтернативный взгляд на проблему дает постановка следующего вопроса: обременено ли поведение в том виде, в каком мы его воспринимаем, ментальными свойствами или нет?
По перцептуализму, мы (по крайней мере иногда) воспринимаем состояния психики в поведении — действие (напр., ходьба по комнате) может быть ощутимо злым или грустным, целенаправленным, пылким, нерешительным и т.д. Цели, желания, мотивы, эмоции, боль или удовольствие, а также многие другие ментальные состояния проявляются в способе действия. Их нельзя сводить к паттернам предрасположенности к поведению (это было бы равнозначно бихевиоризму), и все же они тесно связаны с поведением концептуально, конститутивно или каузально, что обосновывает их доступность для восприятия.
Джеймисон выступает за перцептуализм, подчеркивая, что наша повседневная практика приписывания психических состояний нечеловеческим животным глубоко укоренена, имеет автоматический, концептуально объединяющий и эмпирически действенный характер (Jamieson 1998). Элементы этой же точки зрения можно обнаружить и у ученых, пишущих о когнитивной этологии, и в витгенштейнианском отношении к вопросам о психике животных (напр., Gaita 2003). В последнее время перцептуализм как теорию социального познания (напр., эмпатии) защищали в философии психологии Захави (Zahavi 2011) и Галлахер (Gallagher 2008).
Если верен инференциализм, то если я вижу, например, собаку, вертящуюся передо мной с игрушкой в зубах, виляющую хвостом и глядящую на меня, я могу рассмотреть возможность того, что собака хочет моего внимания, что она радостна и игрива — однако это лишь гипотеза, и если я собираюсь по праву убедиться в этом, то я должен предоставить в ее пользу весомые доводы, основанные на правомерных предпосылках. Напротив, с точки зрения перцептуализма, я буквально вижу (или по крайней мере кажется, что я вижу), что собака хочет моего внимания, что она радостна и игрива.
Обычно считается, что восприятие обосновывает эпистемическую правомерность убежденности, которая может терять силу при определенных условиях — например, если вы выглядываете на улицу и оказывается, что идет дождь, то вы имеете некоторые основания полагать, что идет дождь.
Сложно отказаться от этого допущения, не поддавшись радикальному всеобщему скептицизму, поскольку столь большое количество наших убеждений зиждется на восприятии. Если имеет место кажимость того, что нечто воспринимается, и она обеспечивает то, что имеет место эпистемическая правомерность убежденности в этом, тогда обстоятельство, что я, как кажется, воспринимаю собаку как радостную и игривую, обосновывает правомерность моей убежденности в том, что собака радостная и игривая, то есть обосновывает правомерность моего приписывания собаке ментальных состояний.
С учетом такого prima facie эпистемического подкрепления кажущегося восприятия ментальных состояний у знакомых нам животных, таких как собаки, перцептуалисты склонны полагать, что только колоссальное количество свидетельств могло бы опровергнуть интуитивное приписывание ментальных состояний животным с позиции здравого смысла. Хотя, как обсуждалось выше, Каррутерс (Carruthers 1998) утверждает, что поскольку его теория отрицает наличие сознания у животных, мы должны стремиться искоренить наше интуитивное приписывание сознания, перцептуалист ответил бы на это, что свидетельства, полученные на основе нашего эмпирического соприкосновения с собаками, убедительнее его аргументов (с их опорой сугубо на достоверность его теории высокоуровневого мышления для трактовки сознания; см. разд. 6.1).
Тем не менее, даже если ментальные состояния других людей и известных нам животных, таких как домашние собаки, доступны для нашего восприятия, существуют строгие границы того, насколько далеко это может нас завести при решении общей проблемы наличия сознания у животных. Прежде всего — доступность нашему восприятию может ограничиваться известными нам животными, которые без проблем взаимодействуют с людьми и биологически очень похожи на людей (т.е. млекопитающими).
Во-вторых, как утверждает Деннет, есть примеры, которые показывают, что наше восприятие ментальных состояний можно обмануть. Представляется, будто перцептуализм Деннет оценивает по достоинству, но при этом считает, что восприятие ментальных состояний особенно ненадежно. Тем не менее робот «Cog» (любимый пример Деннета), в отличие от нечеловеческих животных, был специально разработан инженерами так, чтобы выглядеть естественно, то есть чтобы подражать динамическим свойствам движения, запускающим наше восприятие того, что перед нами предмет, наделенный психикой.
Следовательно, причин опасаться, что наше кажущееся восприятие наличия психики у других подрывается подобными примерами, у нас, возможно, не больше, чем причин опасаться, что наше собственное восприятие объектов в пространстве подрывается существованием фотографии.
В обоих случаях инженеры нашли способ создать иллюзию восприятия. Существуют фундаментальные вопросы о том, как восприятие тел в движении может раскрыть ментальное состояние других людей и как вообще возможно визуальное восприятие объектов в пространстве. Однако, вопреки мнению Деннета, нет никаких явных причин, по которым существование тщательно созданных иллюзий подрывало бы общую эпистемическую ценность восприятия.
В-третьих, даже среди тех ученых, кто считает, что они чутко наблюдают за животными с богатой ментальной жизнью, есть те, кто признает, что научный контекст требует от них предоставить определенный вид обоснования эмпирической правомерности приписывания ментальных состояний. Требование подразумевает следующее: говорящий о том, что расхаживающему в зоопарке тигру «скучно» или что рыба, пойманная на крючок, испытывает боль, должен дать определение терминам, указать эмпирический критерий их применения, а также предоставить свидетельства в пользу своих утверждений, основанные на экспериментах или наблюдениях. Даже если перцептуализм является жизнеспособной теорией обыденной практики, связанной с приписыванием сознания животным, представляется маловероятным, что он сможет инкорпорироваться научной эпистемологией.
Познание и сознание
Многие ученые и философы по-прежнему убеждены в том, что даже если некоторые вопросы о психике животных разрешимы эмпирическим путем, никакое количество экспериментов не сможет обеспечить доступ к феноменальному сознанию как таковому. Сторонники такого мнения встречаются даже среди тех, кто готов прибегать к когнитивным объяснениям поведения животных, ссылающимся на внутренние представления. Нежелание иметь дело с сознанием можно трактовать частично как наследие бихевиористской психологии: во-первых, из-за того, что бихевиористы отвергают любые термины для ненаблюдаемого, если их нельзя формально определить через наблюдаемое или же операционализировать экспериментально; во-вторых, из-за сильно укоренившейся в сознании многих бихевиористов ассоциации, связывающей использование менталистских терминов с двойной угрозой картезианского дуализма и интроспективной психологии. В некоторых случаях эти ученые даже придерживаются дуализма, однако они твердо отрицают возможность научного исследования сознания и по-прежнему скептически относятся ко всем попыткам полноценно ввести его в научный оборот.
Кроме того, важный ход аргументации философов заключается в том, что субъективная природа сознания делает его изначально сложным для изучения. Различение Блоком феноменального сознания и сознания доступа (Block 1995), имевшее большое влияние, подавалось как часть критики того подхода к сознанию, который представлен в психологической литературе.
Концептуальное различение феноменальности и доступности, а также сложность эмпирического обоснования сознания доступа рассматривались как главная преграда на пути эмпирических исследований сознания. Сам Блок в последнее время более оптимистично настроен и даже утверждает, что определенные эксперименты могут провести границу между феноменальным сознанием и сознанием доступа (Block 2011). Однако различение феноменального сознания и сознания доступа действительно ставит специфические методологические преграды для тех, кто хочет исследовать сознание доступа эмпирически.
Поскольку предполагается, что сознание приватно или субъективно, как правило, считают, что оно находится за пределами объективных научных методов (напр., Нагель 2003). Это утверждение можно воспринимать двояко. С одной стороны, можно считать, что оно существенно для ответа на вопрос о распределении, то есть позволяет отвергнуть возможность познания того, что представитель другой таксономической группы (напр., летучая мышь) имеет сознательные состояния. С другой стороны, можно считать, что оно существенно для ответа на феноменологический вопрос, то есть позволяет отвергнуть возможность познания феноменологических подробностей психических состояний у представителей иной таксономической группы. Разница между убеждением, имеющим правомерное обоснование, что летучая мышь сознательна, и знанием того, каково быть летучей мышью, важна, поскольку в лучшем случае приватность сознательного опыта подтверждает отрицательный вывод лишь для последнего утверждения.
Чтобы подтвердить отрицательный вывод для первого утверждения, необходимо также предположить, что сознание не оказывает абсолютно никакого измеримого влияния на поведение, то есть необходимо принять эпифеноменализм. А если эпифеноменализм отвергается и заявляется, что сознание в действительности оказывает влияние на поведение, для того чтобы подтвердить применимость этого тезиса, можно прибегнуть к стратегии заключения к наилучшему объяснению. Более того, если определенные сознательные состояния оказывают определенное влияние на поведение, эта стратегия может быть использована для прояснения ряда черт сознательного опыта других животных, даже если некоторые особенности останутся недоступными из-за нашей (как людей) неспособности в полной мере осознать, каково их испытывать. Более подробно об этом будет сказано в следующем разделе.
Если феноменальное сознание полностью эпифеноменально, как полагают некоторые философы, то поиск функций сознания обречен на безрезультатность. Действительно, если сознание полностью эпифеноменально, то оно не могло сформироваться в результате естественного отбора. Исходя из предположения, что феноменальное сознание характерно для человеческой психики, сформировавшейся в процессе эволюции, и следующей отсюда ложности эпифеноменализма можно заключить, что попытка понять биологические функции сознания дает наибольшие шансы зафиксировать его формирование у различных видов. (См. более подробное обсуждение этого вопроса: Robinson 2007.)
Дуализм и физикализм
Пока эпистемологическая и связанная с ней методологическая проблематика находятся на переднем крае дискуссий о сознании у животных, философское рассмотрение сознания в рамках аналитической традиции на протяжении нескольких последних десятилетий фокусируется на метафизических вопросах о природе феноменального сознания и на том, насколько оно вписывается (или не вписывается) в рамки натурализма.
Можно решить, что вопрос о том, что такое сознание, должен быть (метафизически) решен до обращения к вопросу распределения — что онтология должна обуславливать эпистемологию. Однако метафизические вопросы, занимающие аналитических философов последние несколько десятилетий, в значительной степени не связаны с проблемой распределения, больше зависящей от вопросов о структуре и функциях сознания, которые обсуждаются ниже.
Традиционная проблема соотношения ментального и физического касается метафизического статуса психики по отношению к физическому миру (см. статью про дуализм).
Дуалистические теории сознания обычно отрицают то, что его можно объяснить с помощью естественных наук в их наличном виде. Традиционные дуалисты могут утверждать, что к физически описываемым механизмам сознание нельзя сводить (или невозможно свести) при любом понимании физического. Другие могут полагать, что сознание есть все-еще-не-описанная фундаментальная составляющая физического мира, несводимая ни к одному из известных физических принципов. Подобные трактовки сознания не содержат, однако, никаких серьезных причин сомневаться в том, что животные обладают сознанием (возможное исключение представляют те, что основываются на антропоцентричной теологии).
Картезианский дуализм, конечно, традиционно ассоциируется с представлением, что у животных нет разума. Однако аргументы Декарта опирались не на онтологические принципы, а лишь на то, что, с его точки зрения, свидетельствовало о неспособности животных рационально использовать язык или в целом мыслить. На этом основании он утверждал, что нет ничего в поведении животных, что требовало бы объяснения не механистического, а менталистского, и потому не видел смысла приписывать им разум.
Так, картезианский аргумент в пользу исключительности сознания человека отчасти основывается на предпосылке, что для объяснения таких способностей человека, как язык, рациональность и самосознание (осведомленность о себе как о предположительно по сути мыслящей вещи), материальных процессов недостаточно — а следовательно, для объяснения этих явлений использовалась нематериальная душа. Сегодня лишь немногие согласятся с тем, что материальных процессов недостаточно для появления таких сложных явлений, как язык и рациональность.
Действительно, наше понимание «материального» претерпело существенные изменения со времен Декарта. Однако субъективная природа сознания по-прежнему заставляет некоторых авторов утверждать, что психические явления не могут быть сведены к физическим явлениям.
Нет никакой концептуальной причины, по которой тело животного меньше подходит на роль носителя картезианской души (или каких-либо других предположительно нефизических аспектов сознания, постулируемых приверженцами дуализма), чем человеческое тело. Следовательно, в рамках дуализма отрицание наличия сознания у животных не является концептуально необходимым. Для дуалистических, также физикалистских теорий, распределение сознания есть вопрос эмпирической вероятности. Для некоторых дуалистов он может сводиться к вопросу о том, обладают ли животные, о которых идет речь, специфическими познавательными способностями, в то время как другие могут полагать, что нефизическая природа ментального затрудняет или делает невозможным его эмпирическое исследование.
Ранние трактовки сознания в рамках физикализма исследовали философские следствия отождествления сознания с некими физическими или психологическими свойствами нейронов. В такой общей форме подобные теории не содержали никаких особых препятствий для приписывания сознания животным, учитывая то, что в основе животных и людей лежит один и тот же набор биологических, химических и физических принципов. Если можно было бы определить, что феноменальное сознание тождественно или, по меньшей мере, отлично коррелирует с каким-либо общим свойством (как, напр., когерентные квантовые процессы в микротрубочках нейронов или определенная частота мозговых волн), то решение вопроса распределения сводилось бы к непосредственной задаче установить, обладают ли представители других видов этими специфическими свойствами или нет. Даже Серл (Searle 1998), отрицающий диалектику физикализм/дуализм, допускает, что как только нейроученые справятся с нетривиальной задачей определить физиологическую основу сознания у животных (напр., у млекопитающих), при которой не возникало бы никаких обоснованных сомнений в их сознательности, вопрос распределения станет тривиальным, даже в сложных случаях (напр., в случае насекомых).
Нейрофункциональные трактовки
Некоторые философы искали более точное обоснование своих интерпретаций сознания в неврологии. Стратегия Блока (Block 2005) — использовать предварительные функциональные характеристики феноменального сознания и сознания доступа для того, чтобы интерпретировать те свидетельства, которые были получены нейроучеными в процессе поисков нейрокоррелятов сознания. Основываясь на тех свидетельствах, что были получены в ходе экспериментов с людьми и с обезьянами, он утверждает, что наиболее вероятным кандидатом на роль нейрокоррелята феноменального сознания у этих видов является повторяющаяся обратная связь в сенсорной коре. Принц (Prinz 2005) также придерживается нейрофункциональной трактовки, но в отличие от Блока отводит феноменальному сознанию иную функциональную роль. Он высказывается в пользу отождествления феноменального сознания с мозговыми процессами, задействованными при рассмотрении перцептивных представлений среднего уровня, которые способствуют работе памяти посредством высокоуровневых инвариантных представлений.
Тем не менее Принц утверждает, что нам, возможно, никогда не удастся ответить на вопрос распределения в отношении более отдаленных видов — в этом контексте он упоминает осьминогов, голубей, пчел и слизней.
Репрезентативистские трактовки
Репрезентативистские теории сознания связывают феноменальное сознание с репрезентационным содержанием ментальных состояний, соответствующих определенным функциональным критериям.
Репрезентативистские трактовки первого уровня постулируют, что если определенное состояние визуальной системы организма отражает определенное свойство мира функционально подходящим образом (напр., без понятийного опосредования, как часть сенсорной системы), то организм феноменологически сознает это свойство. Трактовки первого уровня обычно вполне совместимы с приписыванием сознания животным: наличие у животных внутренних состояний с необходимыми функциональными и репрезентативными свойствами является относительно непротиворечивым (в той степени, в какой ментальная репрезентация сама по себе является непротиворечивой). Такой подход лежит в основе утверждения Дретцке, что феноменальное сознание неразрывно связано со способностью существа воспринимать и реагировать на особенности его окружения (Dretske 1995), то есть со одним из бесспорных вариантов понимания сознания, который был обозначен выше. Следовательно, с точки зрения Дретцке, феноменальное сознание широко распространено в животном царстве. Сходным образом Тай на основе своей репрезентативистской трактовки феноменального сознания первого порядка утверждает, что оно простирается даже на медоносных пчел (Tye 2000).
Многие философы предложили разнообразные высокоуровневые трактовки феноменального сознания вследствие большого числа предположительно контринтуитивных следствий первоуровневых теорий. Подобные трактовки для объяснения феноменального сознания используют ментальные состояния, направленные непосредственно на другие ментальные состояния.
«Теория высокоуровневого мышления» (ВУМ) Каррутерса состоит в том, что ментальное состояние феноменально сознается субъектом только в том случае, если оно доступно этому субъекту для того, чтобы мыслить его непосредственно (Carruthers 1998a,b, 2000). Термин «доступно» превращает в данном случае эту трактовку в «диспозиционалистскую».
Противоположной будет являться «актуалистская» трактовка, в соответствии с которой субъект может считаться сознательным в соответствующем смысле слова только при актуальном возникновении у него высокоуровневого мышления. По мнению Каррутерса, такое высокоуровневое мышление возможно только в том случае, если существо имеет «теорию психики», обеспечивающую его понятиями, необходимыми для размышления о ментальных состояниях. В современном контексте точка зрения Каррутерса представляет особый интерес, поскольку он использует ее в явном виде для отрицания наличия феноменального сознания у (почти) всех нечеловеческих животных.
Каррутерс утверждает, что научные свидетельства в пользу наличия у нечеловеческих животных теории психики практически отсутствуют (даже у человекообразных приматов; возможное исключение составляют лишь шимпанзе). Из этого он делает вывод, что и мнение, будто любые животные обладают феноменальным сознанием, также не имеет под собой практически никаких оснований. Дальнейшая оценка подобного рода аргументации будет приведена ниже, здесь же стоит отметить, что если (судя по данным экспериментов по приписыванию ложных убеждений) у детей до четырех лет теория психики обычно отсутствует, то позиция Каррутерса подразумевает, что они также не обладают чувствительностью — несмотря на страх перед иглами! Каррутерс стойко переносит это затруднение, хотя для многих оно редуцирует его точку зрения (реакция, которую Каррутерс наверняка бы счел спорной).
В отличие от Каррутерса, рассматривающего чувствительность исходя из высокоуровневого мышления, другие теоретики, такие как Армстронг и Ликан, отдают предпочтение трактовке с точки зрения высокоуровневого опыта, в рамках которой сознание объясняется через внутреннее восприятие ментальных состояний (Armstrong 1980, Lycan 1996). Истоки ее восходят к Аристотелю, а также к Джону Локку. Поскольку такие модели не требуют способности концептуализировать ментальные состояния, сторонники теорий высокоуровневого опыта чаще склонны допускать, что такие способности могут быть обнаружены у других животных, нежели сторонники теории высокоуровневого мышления [6]. Однако Дженнаро утверждает, что теория высокоуровневого мышления совместима с признанием наличия у нечеловеческих животных сознания. Он полагает, что Каррутерс и другие преувеличивают требования к тому, какие ментальные понятия являются необходимыми, и что сеть проводящих путей головного мозга животных обеспечивает структуру, в которой представления более высокого и более низкого уровня действительно могут объединяться в рамках единого сознательного состояния (Gennaro 2004).
Бинарно ли сознание?
Один из метафизических вопросов, непосредственно относящийся к вопросу о филогенетическом распределении и эволюции сознания, заключается в том, может ли обладание сознанием (состояние сознательности) быть описано как бинарное (имеется / отсутствует) или же у него имеются градации. Некоторые авторы, причем абсолютно по разным причинам, отрицают общее, но с их точки зрения проблематичное допущение, что «сознание есть переключение между вкл/выкл; система либо сознательна, либо нет» — так это представление формулирует Серл, вполне разделяющий его (приводится по Lycan 1996, где данный тезис отвергается).
Ликан утверждает: существует широкий спектр степеней богатства или полноты сознания; существуют имеющие большое значение случаи, когда система с минимальной степенью сознания не является «действительно» сознательной (Lycan 1996: 8). Конечно, звучит это несколько парадоксально, но суть, по всей видимости, в том, что признавать крайне низкие уровни сознания для нас контринтуитивно, поскольку сложно представить себе содержание очень простых ментальных состояний. Одно из толкований заключается в том, что с точки зрения Ликана «сознательный» — неопределенный, или смутный (vague), предикат, а сознание не распределено линейным образом.
Деннет также считает, что сознание не является бинарным. Он утверждает это, выступая в защиту радикально дефляционного антиреализма в отношении сознания в целом, в соответствии с которым сознание — фактически иллюзия, порожденная языком (Dennett 1991/1995).
Велманс различает «теории прерывности», постулирующие существование определенного момента возникновения сознание, до которого оно не существовало (что относится ко вселенной в целом и к любому конкретному индивидуальному сознанию), и «теории непрерывности», рассматривающие эволюцию сознания с точки зрения «постепенного перехода сознания от нераспознаваемого к распознаваемому» (Velmans 2012). С его точки зрения, теории непрерывности более элегантны, так как любая прерывность обусловлена произвольным критерием, а «трудная проблема» стоит перед ними гораздо острее, чем перед теориями преемственности.
Три автора, чьи точки зрения описаны выше, отрицают бинарность сознания исходя из очень разных оснований, и каждая из этих точек зрения содержит противоречия. Более того, ни одна из них особенно не предлагает инструментов или идей для понимания того, как мы можем мыслить предположительно небинарную природу сознания. Исходя из тезиса Ликана, касающегося наличия степеней богатства или полноты, можно задаться вопросом, какие градации измерений или качественных параметров доступны для различения различных видов психики. Многие авторы различают «первичное» и «высокоуровневое» сознание (Seth et al. 2005); «первичное», «вторичное» и «третичное» сознание (Panksepp 2005); «основное» и «расширенное» сознание (Damasio 1999).
Однако кажется, что большинство из них соотносит феноменальное сознание (т.е. обладание каким бы то ни было субъективным опытом) с первичным или основным сознанием. Предполагается, что термины «вторичный» и «третичный» указывают на развитые формы сознания. В этой связи неясно, противоречит ли какая-либо из этих классификаций представлению о том, что феноменальное сознание само по себе является бинарным, что оно либо целиком имеется, либо целиком отсутствует у данной системы. Это затруднение заслуживает более пристального внимания, поскольку оно затрагивает проблемы распределения и эволюционного происхождения сознания. Если сознание не является бинарным, то его распределение не будет иметь строгих границ, а будет включать градации, то есть некоторые животные могут быть частично или неполностью сознательными.
Ограничения философских теорий
Феноменальное сознание — всего лишь одна из черт (определяющая, как сказали бы некоторые), характеризующих ментальные состояния или события. Однако любая теория сознания животных должна рассматриваться в контексте более широкого исследования познавательных способностей животных, которое также будет затрагивать (среди философов) вопросы интенциональности (в том смысле, в каком она описывается немецким психологом XIX века Францем Брентано) и ментального содержания (Dennett 1983, 1987; Allen 1992a,b).
Среди философов имеются разногласия по вопросу отношения сознания к интенциональности: одни утверждают, что сознание и интенциональность строго независимы; вторые (в частности, сторонники описанных здесь функционалистских теорий сознания) полагают, что интенциональность необходима для сознания; третьи считают, что сознание необходимо для подлинной интенциональности. Многие ученые, исследующие поведение, принимают когнитивистские объяснения поведения животных и приписывают предметам своего исследования репрезентативные состояния. Однако приписывать сознание они не решаются.
Что касается метафизики феноменального сознания и его точном соотношении с интенциональностью, с мозгом, с поведением и т.д., в этих вопросах сохраняется большая неопределенность. Исследование основательной критики различных трактовок сознания, использующих эти термины, выходит за рамки настоящей статьи, однако можно с уверенностью сказать, что ни одно из направлений этой критики не выглядит достаточно надежным, чтобы положить конец спорам о наличии или отсутствии сознания у животных. Интерпретации сознания в терминах основных нейрофизиологических свойств, квантово-механических свойств нейронов, или sui generis свойств вселенной столь же ненадежны, как и различные функционалистские трактовки. И даже те интерпретации, которые в своем общем виде совместимы с животными, не достаточно конкретны и не могут дать готовые ответы на вопрос распределения во всей его полноте. Следовательно, на основе обсуждавшихся до сих пор философских теорий сознания нельзя сделать никаких однозначных выводов о распределении сознания.
Где в таком случае остаются эпистемологические вопросы о сознании животных? Убежденность, что мы должны иметь теорию о том, что такое сознание, прежде чем мы попытаемся определить, есть ли оно у других животных, может казаться естественной, но на самом деле в этом случае мы ставим концептуальную телегу впереди эмпирической лошади. На ранних этапах научного исследования любого явления предполагаемые образцы должны определяться в соответствии с приблизительными правилами из опыта (или с рабочими определениями), а не в соответствии с полноценными теориями. Древние ученые определяли золото по случайным характеристикам, а не в соответствии с его атомарной сущностью — этого знания пришлось дожидаться и тщательно исследовать многие предполагаемые образцы, некоторые из которых оказались золотом, а некоторые нет. Сходным образом, на данном этапе игры изучение сознания животных возможно выиграло бы, если мы определим признаки животных, достойных дальнейшего изучения, не будучи твердо уверены в том, что все выбранные образцы окажутся связаны с сознательным опытом.
Вспомним аргумент Декарта о том, что животные не используют для разговора язык или разум в целом. Этот аргумент, основанный на предполагаемой неспособности животных проявлять определенные интеллектуальные способности, иллюстрирует общую закономерность использования определенных различий между животными и людьми для утверждения того, что у животных отсутствует сознание.
Обычный рефрен, который мы слышим в ответ на подобные доводы, таков: в условиях неполной информации «отсутствие доказательств не доказывает отсутствия». Декарт сбрасывал со счетов попугаев, произносящих человеческие слова, потому что считал их просто лишенным смысла повторением. Возможно, убеждение было оправданно в случае тех немногочисленных попугаев, с которыми он имел дело, однако оно не было основано на систематическом научном изучении способностей попугаев. В настоящее время многие утверждают, что исследование Пепперберг африканского серого попугая по имени Алекс должно похоронить это картезианское предубеждение (Pepperberg 1999). Наряду с несколькими исследованиями приобретаемой степени коммуникативной компетентности у шимпанзе и бонобо (напр., Gardner et al. 1989; Savage-Rumbaugh 1996) данное исследование, похоже, подрывает утверждения Декарта о том, что у животных отсутствует смысловая коммуникация и общие способности размышления. (См. также статьи в Hurley & Nudds 2006.)
Картезианцы в ответ обращают внимание на те ограничения, которые демонстрируют животные в таких исследованиях (ведь они не могут хорошо играть в шахматы, не говоря уже о том, что не могут сказать нам, о чем думают), и присоединяясь к лингвистам, возмущаются, что объекты исследований по изучению языка животных не овладели рекурсивным синтаксисом естественных человеческих языков в полной мере [7]. Однако подобное ретроспективное повышение планки демонстрирует многим ученым, что картезианская позиция — не научная гипотеза, а догма, которую необходимо защищать любыми средствами.
Сегодня свидетельства о наличии у животных сложных познавательных способностей убедительны для большинства философов, и они согласны с Блоком.
Ведь, несомненно, большинство согласится с тем, что на определенном этапе различия между способностями человека и представителями другого вида (напр. Lumbricus terrestris, обыкновенного земляного червя) столь велики, что едва ли такие существа обладают чувствительностью.
Неоднозначность возникает именно потому, что никто не может сказать, какую степень различия считать достаточной для констатирования отсутствия чувствительности.
Стремление осуществить последовательный отбор тех поведенческих или нейрофизиологических характеристик, которые могли бы служить надежными индикаторами наличия сознания, стимулирует развитие подхода, ориентированного на структуру и функционирование, который применялся многими авторами с начала XXI века. Хотя иногда этот подход сопровождался метафизическими вопросами о сознании, данный проект дает надежду на эмпирическую трактовку даже в свете сохраняющейся неопределенности по метафизическим вопросам, касающимся сознания.
Структура и функционирование сознания
Одной из стратегий введения сознания в научный оборот является попытка сформулировать теоретическое основание для соединения наблюдаемых характеристик животных (поведенческих или неврологических) и сознания. Какое влияние сознание должно оказывать на поведение? Наличие каких способностей и настроений мы должны ожидать у сознательного существа, которые могут отсутствовать у существа, не обладающего сознанием? Какие нейрофизиологические структуры и процессы могли бы реализовать динамику или обработку информации, которые необходимы для сознания?
Подобный подход возникает у Гриффина, который пытается заставить этологов обратить внимание на вопросы сознания у животных, причем во всех смыслах этого термина — включая феноменальное сознание.
Гриффин, снискавший свою научную репутацию благодаря тщательному описанию физических и физиологических характеристик эхолокации у летучих мышей, в сери книг приводит примеры наличия у животных коммуникативного поведения, а также поведения, направленного на решение задач, которое особенно проявляется в природных условиях — и утверждает, что именно в таких условиях ученые-этологи должны начинать свои исследования сознания животных (Griffin 1976, 1984, 1992).
Хотя Гриффин считает, что демонстрируемый в этих примерах интеллект предполагает наличие сознательного мышления, многие критики были разочарованы отсутствием последовательной связи между его примерами и приписыванием сознания (см., напр., Alcock 1992). Основным конкретное предположением Гриффина в этом отношении было довольно неправдоподобно — будто сознание может функционировать как компенсация ограниченной нейронной системы. Таким образом, Гриффин выдвигает предположение, что сознание может быть более важным для медовых пчел, чем для людей.
Если компенсация в отношении небольших наборов нейронов не является правдоподобной функцией сознания, то что же может выступить в этом качестве? Ответ с позиций здравого смысла звучал бы так: сознание «рассказывает» организму о событиях в окружающей среде или, в случае боли и других проприоцептивных ощущений, о состоянии тела.
Однако такой ответ не учитывает различие между феноменальным сознанием и простой осведомленностью (в смысле обнаружения, который является бесспорным) о событиях окружающей среды или телесных событиях, и потому порождает вопросы в адрес противников приписывания животным сознательных состояний. Противники приписывания животным феноменального сознания не отрицают существование более общего вида сознавания различных внешних и телесных событий, а значит нет никакого логического перехода от наличия осведомленности о вещах в окружающей среде или в теле к наличию чувствительности у животных.
Возможно, более изощренные попытки прояснить функционирование сознания также обречены на неудачу. Однако Аллен и Бекофф предполагают, что изучение способностей животных корректировать свои собственные ошибки восприятия может продвинуть нас вперед (Allen & Bekoff 1997: ch. 8). Не все корректировки ошибок дают основания подозревать, что в этот процесс вовлечено сознание, однако в тех случаях, когда организм может скорректировать ошибку восприятия, сохраняя при этом способность использовать содержание ошибочного восприятия, может возникнуть устойчивое впечатление, что животное внутренне отличает состояния собственной явленности от других суждений о мире.
(Люди, напр., имеют сознательный зрительный опыт и знают о том, что в некоторых случаях он вводит их в заблуждение — т.е. что существуют зрительные иллюзии, — однако они могут использовать ошибочное содержание такого опыта в различных целях, в частности, чтобы вводить в заблуждение других или отвечать на вопросы о том, как им являются эти вещи.) Учитывая, что существуют теоретические основания для отождествления сознательного опыта с «состояниями явленности», возможно, стоит начинать искать сознание у животных именно через попытки выяснить, обладают ли они такими способностями.
Однако важно подчеркнуть, что такие способности сами по себе не должны рассматриваться как характерные или определяющие для сознания. Каррутерс делает аналогичное предположение о функционировании сознания, связывая его с общей способностью проводить различие между явлением и реальностью; разумеется, он продолжает утверждать, что эта способность зависит от наличия концептуальных ресурсов, которые недоступны нечеловеческим животным (Carruthers 2000).
Обширная проблематика, касающаяся функционирования, тесно связана с вопросами о том, ментальный процесс какого сорта представляет собой сознание.
· Структура. Что выступает содержанием сознания (какую информацию, представления, интенциональное содержание, свойства, процессы и т.д. оно включает)? Какая (возможно, бессознательная/подсознательная) информация, какие представления или другие когнитивные и интенциональные процессы, сущности и отношения необходимы для сознания?
· Функционирование. Как сознание соотносится с другими (неосознанными) процессами в познании, теле и окружающей среде? Что привносит обладание сознанием в способность животного ориентироваться и адаптироваться к окружающей среде, выживать и процветать?
Биологически ориентированные представления о том, какие животные обладают сознанием, когда, как и почему оно сформировалось, в значительной степени зависят от выбора той или иной точки зрения по следующим вопросам: что представляет собой сознание как когнитивный процесс и как оно связано с другими биологическими процессами (поведением, развитием и экологическим взаимодействием). Проиллюстрируем тезис на примере грубого различения двух точек зрения.
Некоторые рассматривают сознание как нечто основополагающее для базовых перцептивных и когнитивных процессов, вовлеченных в управление телом животного.
Другие рассматривают сознание как нечто добавочное или подключаемое к системе, которая уже является достаточной для базового контроля действий, управляемых восприятием. Чем более основополагающим является сознание для базового функционирования животных, тем более широко оно должно быть распространено и тем более древним оно должно являться.
Однако в случае, если сознание представляет собой явление относительно модульное, функционально узкое и концептуально высокоуровневое, то мы должно считать, что оно ограниченно распределено среди животных и возникло относительно недавно. Все взгляды, которые рассматриваются в следующем разделе, разрабатывают связь между функционированием, структурой, распределением и эволюционным происхождением сознания.
Необходимо отметить еще один момент: правомерность структурных моделей сознания обычно обосновывается феноменологическими или интроспективными наблюдениями, т.е. наблюдениями о природе сознания в том виде, в каком оно является частью опыта субъекта.
Хотя на протяжении всего XX и XXI веков использование подобных методов от первого лица в психологии и философии являлось спорным, в настоящее время, похоже, распространенным является признание их неотъемлемой роли в научном исследовании сознания, и многие авторы, недавно опубликовавшие свои научные теории сознания, периодически используют феноменологические предпосылки для обоснования своих взглядов (напр., Seth, Edelman and Baars 2005; Merker 2005; Tononi 2008; Cabanac et al. 2009).
Эволюция и распределение сознания
Ученые и философы выдвинули множество гипотез о том, какие животные обладают сознанием, а какие нет. Эти взгляды охватывают огромный спектр возможностей, от самых ограниченных — что сознанием обладают только люди, до самых широких –некоторые авторы утверждают, что почти все животные, даже простые беспозвоночные, обладают базовой способностью переживать мир в своем опыте. Часть авторов даже утверждает, что одноклеточные организмы (Margulis 2001) или растения (A. Nagel 1997) являются сознательными, а некоторые приводят аргументы в пользу тех или иных версий панпсихизма, представления о том, что сознание является свойством фундаментальных физических сущностей, практически так же, как масса и заряд (Chalmers 2015).
Следует отметить, что ни приписывание сознания одноклеточным организмам, ни фундаментальным физическим сущностям не подразумевает, что все животные обладают сознанием.
В первом случае может оказаться, что сложность обработки информации и объединение относительно сложных одноклеточных организмов превосходит возможности простейших животных. Во втором случае, хотя в разработанной Чалмерсом версии пансихизма «микроопыт» приписывается «фундаментальным физическим сущностям», отсюда не следует, что какой-либо особый макроскопический объект (как, напр., животное) имеет «макроопыт» — тот «вид сознательного опыта, которым обладает человек» (Chalmers 2015).
Далее мы не будем обсуждать эти вопросы, поскольку они выходят за рамки темы настоящей статьи.
Вопрос о том, какие родственные группы (виды, или группы более высокого порядка, такие как класс или тип) животных являются сознательными, неминуемо сопровождается рассмотрением вопроса об эволюционном происхождении сознания.
Это логическое следствие дарвинистского (в широком смысле слова) понимания жизни, в соответствии с которым современные организмы произошли видоизменяясь, от небольшого количества (возможно, от одного) очень древних предков.
Если какая-либо черта присуща определенному виду, то она либо возникла у представителей этого вида, либо происходит от предка — и в этом случае она будет присуща и другим видам, произошедшим от того же предка, если только она не превратилась во вторичный признак.
Возникло ли сознание впервые у человека или у более раннего нечеловеческого предка? Если верно последнее, то какой же предок это был? Возможно также, что сознание возникало несколько раз, как способность летать при помощи крыльев, которая появилась в процессе эволюции у насекомых, птиц, летучих мышей и птерозавров независимым образом.
Люди
Как было описано выше, точка зрения, в соответствии с которой сознание присуще исключительно человеку, имеет долгую историю. Она согласуется с религиозным взглядом на человечество как вершину мироздания, а также может подкупать, поскольку освобождает нас от всякой вины за наше обращение с животными. Эта точка зрения может получить не только религиозную, но и значительную интуитивную поддержку, поскольку заманчиво соединить сознание с проблемой человеческой уникальности. Если сознание можно было бы связать с языком, абстрактным мышлением или какой-то другой ментальной характеристикой, потенциально объясняющей нашу явную обособленность от природного мира, то это решило бы сразу две неординарные загадки.
Деннет выступает как решительный сторонник представления о том, что человек является уникальным обладателем сознания (Dennett 1991, 1995). Деннет отстаивает эту позицию вкупе со своей антиреалистической теорией сознания, основным пунктом которой является положение о том, что сознание есть своего рода «иллюзия пользователя» (Dennett 1995) или «вымысел» (Dennett 1991: 365: 429), построенным людьми посредством нарративных описаний:
То, что существует на самом деле, есть просто различные события фиксации содержаний, происходящие в мозгу в разное время и в разных местах… Некоторые из этих фиксаций содержаний имеют дальнейшие следствия, которые в конечном итоге приводят к высказыванию предложений — на естественном языке — публично или только внутренне. И так создается гетерофеноменологический текст… А что же настоящая феноменология? Такой вещи не существует. (Dennett 1991: 365)
С точки зрения Деннета, только люди (взрослые, хорошо встроенные в культуру и обладающие языковой способностью) обладают сознанием, поскольку сознание есть разновидность повествования, что требует языка, а только люди владеют языком.
Каррутерс также отстаивает точку зрения, в соответствии с которой только люди обладают сознанием (возможное исключение представляют шимпанзе), однако исходя из оснований, отличных от аргументов Деннета. Каррутерс приводит доводы, основанные на его теории «высокоуровневого мышления», согласно которой феноменальное сознание требует способности размышлять о своих собственных мыслях, и следовательно концептуализировать их [8]. Подобная концептуализация требует, по мнению Каррутерса, теории психики (Carruthers 1998a,b, 2000).
С точки зрения Каррутерса, у нас мало оснований считать, что у какого-то нечеловеческого животного имеется теория психики; впрочем, возможное исключение составляют шимпанзе (см. подробное обсуждение теории психики у нечеловеческих животных: Lurz 2011, Andrews 2012). Этот аргумент, конечно же, не более силен в сравнении с трактовкой сознания в терминах теории высокоуровневого мышления, на которой он основывается. Однако если закрыть на это глаза из дискуссионных соображений, то данный вызов Каррутерса заслуживает более пристального внимания.
Каррутерс не поддерживает, но и не отвергает напрямую вывод о том, что шимпанзе обладают чувствительностью. В основе его подозрений, что даже шимпанзе могут не обладать теорией психики, а следовательно (с его точки зрения) — и феноменальным сознанием, лежат результаты затейливых лабораторных исследованиях, проведенных Повинелли (Povinelli 1996). Исследования демонстрируют: при взаимодействии с людьми, приносящими им пищу, шимпанзе, по всей видимости, не понимают ту роль, какую глаза играют для человека при получении зрительной информации, хотя их поведение в том, что касается внимания к таким сигналам, как физиогномические данные, внешне сходно с поведением человека. Интерпретация исследования Повинелли остается спорной. Хейр и др. провели эксперименты, в рамках которых доминирующие и подчиненные животные конкурировали друг с другом за пищу, и пришли к выводу:
…по крайней мере в некоторых ситуациях шимпанзе знают, что видят, а что не видят их сородичи, и, более того, используют эти знания, чтобы сформировать собственные поведенческие стратегии в ситуации конкуренции за пищу. (Hare et al. 2000).
Они полагают, что отрицательные результаты, полученные Повинелли, могут объясняться тем, что в его экспериментах имеет место менее естественное для шимпанзе взаимодействие с человеком. На фоне имеющейся двусмысленности Каррутерс высказывается довольно осмотрительно и выдвигает свои тезисы о наличии у шимпанзе чувствительности только в качестве гипотезы.
Полноценный обзор полемики по поводу теории психики (напр., Heyes 1998; Lurz 2011; Andrews 2012) заслуживает отдельной статьи, здесь же стоит упомянуть, что истоком споров выступает гипотеза, согласно которой интеллект приматов в целом и человеческий интеллект в частности специально приспособлены для социального познания (см. Byrne & Whiten 1988, особ. Jolly & Humphrey [ch. 1–2]).
В связи с этим утверждается, что доказательства возможности приписывания ментальных состояний большому количеству видов лучше искать в естественной активности животных (к примеру, в их социальных играх), а не в лабораторных экспериментах, где животные помещаются в искусственные ситуации (Allen & Bekoff 1997 [esp. ch. 6]; см. также Hare et al. 2000, Hare et al. 2001, Hare & Wrangham 2002). Альтернативные подходы, пытавшиеся представить убедительные доказательства наличия теории психики у нечеловеческих животных, рассматриваемых в естественных условиях обитания, в целом не справились с этой задачей (см., напр., выводы о теории разума у верветок и павианов: Cheney & Seyfarth 1990, 2007), хотя устные свидетельства обнадеживают тем, что исследователи просто до сих пор не сумели разработать подходящие эксперименты.
Кроме того, теория психики — и в более широком смысле социального познания — представляет собой область активных исследований, и вполне возможно, что новые работы обнаружат доказательства наличия теории психики у нечеловеческих животных.
По мнению некоторых исследователей, в том числе Каррутерса, сознание основано на когнитивных процессах, крайне специфических и модульных — они в сущности не играют особой роли в процессах восприятия, мотивации и познания, которые задействованы в любом поведении нечеловеческих животных.
Большинство когнитивных процессов (и соответствующих мозговых систем), вовлеченных в человеческую деятельность, имеются и у нечеловеческих животных — согласно этой логике, бόльшая часть человеческой деятельности также является несознательной.
Повреждение зрительной коры делает их феноменологически слепыми в определенных участках их поля зрения («скотомах»), но тем не менее они способны идентифицировать предметы, находящиеся в этих слепых участках. (Оба случая рассматриваются как примеры «бессознательного опыта»: Carruthers 1989, 1992.)
Такое сопоставление поведения животных с бессознательными способностями человека можно критиковать на том основании, что оно опирается лишь на несистематическое наблюдение за поведением животных, как это было в случае высказываний Декарта о попугаях. Есть основания полагать, что тщательное исследование выявит отсутствие значительного сходства между поведением животных и поведением человека, которое постулируется в этих предполагаемых случаях бессознательного опыта.
Примечательно, в частности, что бессознательный опыт автоматического вождения не запоминается субъектом этого опыта, однако нет свидетельств того, что животные так же не могли бы вспомнить свой якобы бессознательный опыт. Аналогичным образом, пациенты со слепозрением не реагируют на предметы, находящиеся в скотомах, спонтанно — они должны быть обучены реагировать при помощи «вынужденной реакции». Нет никаких свидетельств, что подобные ограничения являются нормой для животных или что животные в отношении зрительного опыта ведут себя подобно пациентам со слепозрением (Jamieson & Bekoff 1992).
Тем не менее есть эмпирические основания полагать, что поведение, предполагающее наличие сознания у животных, может быть результатом бессознательных процессов. В одном исследовании рассматривается спинной мозг крыс и их способность к обучению — в нем обнаруживаются явления, аналогичные латентному торможению и затемнению (Allen et al. 2009). Утверждается, что у животных, не подвергшихся экспериментальному воздействию, эти явления, связанные с обучением и памятью, задействуют внимание.
Однако их сходство с процессами, происходящими в спинном мозге (которые по признанию большинства не включают сознание), ставит под сомнение то, что их можно рассматривать как свидетельство в пользу наличия сознания. Конечно, способность спинного мозга к обучению отличается от способности к обучению, имеющейся у не подвергшихся экспериментальному воздействию организмов, и при отсутствии опровержения, имеющиеся доказательства позволяют считать, что сложные формы обучения сопряжены с сознанием (Clark & Squire 1998; Allen 2004; Ginsburg & Jablonka 2007b). Однако это положение сходно с тем, которое было озвучено в отношении слепозрения: выводам о сознании должен предшествовать более скрупулезный анализ таких сходств и различий.
Высшие приматы
Гэллап разработал эксперимент по распознаванию себя в зеркале («зеркальный тест»; Gallup 1970), который стал широко использоваться как тест на осведомленность о себе, хотя интерпретация этого эксперимента остается спорной (см. ниже раздел о самосознании и метакогнициях).
Предвосхищая Каррутерса, Гэллап утверждает, что наличие осведомленности о себе — в смысле способности размышлять о собственных ментальных состояниях — необходимый признак наличия психики, а потому животные, которые «проваливают» зеркальный тест, психики не имеют (Gallup 1982, 1985).
Были споры по поводу того, какие именно животные обоснованно «проходят» те версии теста, которые были модифицированы для использования в экспериментах со слонами, дельфинами и сороками, однако по состоянию на 2002 год Гэллап все еще утверждал, что есть убедительные доказательства того, что люди, обычные шимпанзе, бонобо и орангутанги систематически проходят тест, тогда как многие другие приматы систематически проваливают его. Говорит это о том, с точки зрения Гэллапа, что осведомленность о себе присуща исключительно высшим приматам (Gallup et al. 2002). В сочетании с более ранним тезисом, что сознание не может не подразумевать осведомленности о себе, которая устанавливается при помощи зеркального теста, это положение предполагает, что сознание присуще исключительно высшим приматам.
Гэллаповская интепретация зеркального теста не бесспорна (Mitchell 2002). Рочат и Захави видят в ней сразу два слабых или сомнительных места: (а) интерпретация поведения шимпанзе, ориентированного на зеркало, как индикатора наличия у них опыта зеркального распознавания себя, сходного с человеческим; (б) заявление, что из распознавания себя в зеркале следует наличие сознания (Rochat & Zahavi 2010).
Отметим на полях, что с начала 1990-х годов ведутся споры (Cavalieri & Singer 1994), могут ли высшие приматы рассчитывать на особую правовую защиту, сопоставимую с «правами человека».
Суть дискуссии не в том, имеется ли у высших приматов сознание как таковое (по-видимому, это признает большинство участников дискуссии с обеих сторон), а в том, имеется ли у них личностность. Личностность — понятие проблематичное, как правило, связано оно с определенными формами агентности и осведомленности о себе. Зачастую предполагают, что личностность тесно связана с моральным статусом, что отражает рассматриваемая дискуссия (DeGrazia 1997; Varner 2012; см. также статью о моральном статусе животных). Хотя личностность и не является чем-то существенным для феноменального сознания, нередко считают, что она предполагает наличие сознания и лучше всего мыслить ее как некий уровень развития или сложности сознания.
Млекопитающие
Были выдвинуты разнообразные теоретические и эмпирические аргументы в пользу того, что сознание распределено среди всех млекопитающих. Сет, Баарс и Эдельман утверждают, что нейронные процессы, необходимые для человеческого сознания — распространенная повторяющаяся активность в таламокортикальном комплексе — задействуют анатомические системы, имеющиеся у всех млекопитающих (а может, и у других классов; см. Seth, Baars & Edelman 2005). Панксепп использует аналогичный подход, хотя он фокусируется на нейрофизиологических системах, задействованных в «основных эмоциях». Сторонники обоих вышеупомянутых подходов признают, что сознание может быть распространено шире, не только среди млекопитающих.
Кроме того, необходимо помнить о том, что все последующие предположения подразумевают, что сознание широко распределено среди млекопитающих.
Следовательно, точка зрения, в соответствии с которой все млекопитающие являются сознательными, разделяется множеством ученых, высказывающихся о распределении сознания.
Амниоты (в том числе птицы и рептилии)
Вопросы о том, обладают ли рептилии сознанием (и если да, то какой может быть их психическая жизнь), особенно интересны, поскольку птицы гораздо ближе к рептилиям, нежели к млекопитающим, при этом они демонстрируют такое разнообразие форм поведения, что наблюдатель склонен интуитивно истолковывать их как признаки наличия интеллекта и эмоций, что в случае поведения чешуйчатых, так называемых «холоднокровных» животных (таких как змеи и черепахи), гораздо менее очевидно.
Имеют ли птицы и млекопитающие общие ментальные характеристики (сознание, интеллект, эмоции, социальная привязанность), которые отсутствуют у рептилий? Если да, то это будет являться примером независимой, конвергентной эволюции подобных явлений. Или же такие характеристики являются общими для всех указанных животных, но у одних они выражены менее очевидно, чем у других?
Такое мнение озвучили Канабак с соавторами. Амниоты — это группа, которая включает всех потомков общего предка ныне живущих птиц и млекопитающих, рептилий (ящериц, змей, черепах) и вымерших животных (динозавров, птерозавров и плезиозавров).
В соответствии с гипотезой, только эти животные — в отличие от земноводных, рыб или каких-либо беспозвоночных — сознательны. В основании аргументации лежит сформулированная в явном виде структурно-функциональная теория, трактующая сознание как единое репрезентативное пространство, «абстрактную приватную модель реальности с четырьмя измерениями: качество, интенсивность, гедоничность и длительность» (Cabanac et al. 2009: 268).
Обладание такой способностью моделировать реальность позволяет животным инсценировать возможные образы действия, в качестве «единой валюты» для оценки и выбора действий исходя из ожидаемых последствий (что основано на предыдущем опыте) используется гедонистичность (удовольствие или боль).
На базе данной модели структурно-функциональной теории, Кабанак определяет набор поведенческих индикаторов сознания:
- способность идти на мотивированные уступки;
- игра;
- обход препятствий (что заставляет животное добиваться ряда промежуточных целей безрезультатно, для того чтобы достичь окончательного результата);
- выражение эмоций;
- выражение сенсорного удовольствия;
- эмоциональная лихорадка (повышение температуры тела в ответ на предположительно стрессовую ситуацию, в экспериментах Кабанака — на прикосновения);
- вкусовое отвращение.
Основываясь на предполагаемых доказательствах того, что у амниот имеются эти показатели, а у неамниот — отсутствуют, Кабанак утверждает, что сознание зародилось у общего предка амниот, а значит присутствует у всех ныне живущих амниот, но не у других животных. Кабанак и Кабанак (Cabanac & Cabanac 2009) также утверждают, что качественное отличие роли дофамина в мотивационных процессах в мозге амниот (в сравнении с неманиотами) свидетельствует в пользу этой гипотезы распределения и происхождения сознания.
Кабанак и его коллеги задокументировали наличие некоторых из этих явлений –эмоциональной лихорадки (Cabanac and Bernieri 2000; Cabanac and Cabanac 2000; Cabanac and Cabanac 2004) и вкусового отвращения (Paradis and Cabanac 2004) — у амниот и их отсутствие как минимум у небольшого числа неамниотических видов.
Однако утверждение, что эти аспекты поведения и познания не существуют за пределами группы амниот основано в основном на отсутствии свидетельств в пользу обратного (и, следовательно, оно изначально имеет определенные ограничения). В частности, авторы не предоставляют прямого подтверждения своим заявлениям о том, что неамниоты неспособны идти на уступки ввиду наказаний или вознаграждений, неспособны к игре или к обходу препятствий.
Действительно, некоторые из этих заявлений опровергаются существующими исследованиями — например, описание обхода препятствий у пауков-скакунов (Jackson and Wilcox 2003), а также исследование Элвуда и Аппеля (Elwood & Appel 2009), демонстрирующее мотивированные уступки в поведении крабов-отшельников.
Структурно-функциональную теорию сознания Кабанака можно оценивать независимо от тех свидетельств, которые он собирает в поддержку своего представления о распределении и происхождении сознания.
Действительно, его взгляды на распределение и происхождение можно оспорить как раз принимая его структурно-функциональную теорию и полагая, что предлагаемый им перечень индикаторов сознания может фактически означать более широкое распространение сознания (т.е. если мотивированные уступки, игра и обход препятствий встречаются не только у амниот). Как мы увидим, его представление о структуре и функционировании сознание имеют много общего с представлениями других авторов, выступающих в поддержку тезиса о более широком распределении сознания среди животных.
6.5 Позвоночные
«Рыба» — это термин из народной биологии, который не соответствует в полной мере какой-либо монофилетической таксономической группе. Это можно уяснить, если заметить, что целакантообразные ближе к человеку, нежели к тунцам, или что тунецы ближе к человеку, нежели к акулам.
Это означает, что некоторые создания, которых мы интуитивно причисляем к рыбам, ближе к не-рыбам, нежели к другим рыбам. Как правило, обыденный термин «рыба» относится ко всем позвоночным, кроме четвероногих, хотя он оказывается несколько двусмысленным в отношении таких животных, как морские коньки, угреобразные, миксинообразные и орляковидные.
Так или иначе, имеет место оживленная дискуссия о наличии сознания у рыб, которая в основном сосредоточена на вопросе, может ли рыба испытывать боль, стресс и страдания (см. ниже разд. 7.1; противоположные взгляды на наличие осознанной боли у рыб: Rose 2002, Sneddon et al. 2003; обзоры на тему познания и сознания у рыб и их возможной связи с болью: Allen 2013, Brown 2015; и монографическое исследование: Braithwaite 2010). Он особенно релевантен в контексте регулирования условий работы коммерческих рыбоводств и любительского рыболовства. Соответственно, литература, посвященная наличию сознания у рыб, сосредоточена вокруг опытов и экспериментов, проводимых на семействе лососевых (особенно на лососе и форели). Лососевые имеют большое значение для коммерческого и спортивного рыболовства, но представляют очень незначительную филогенетическую группу тех животных, которых для простоты речи причисляют к рыбам.
Меркер полагает, что сознание зародилось на раннем этапе эволюции позвоночных, и поэтому является одновременно и древним, и широко распространенным (Merker 2005).
В соответствии с подобным предположением, сознанием обладают не только млекопитающие и птицы, но и земноводные, а также все водные позвоночные. Меркер начинает свою аргументацию с феноменологического наблюдения, что содержания сознательного опыта являются объектно- и целеориентированными, но не включают мелкие сенсорные и моторные детали, имеющиеся при периферических и низкоуровневых нейронных процессах по обработке данных. Меркер утверждает, что сознание является объединенной репрезентационной платформой — он называет это «синтезированным пространством реальности», — которая в случае животных со сложноустроенными телами, обеспечивающими множество степеней свободы движения и многочисленные сенсорные модальности, решает целый пласт принципиальных проблем нейронной логистики. В их числе:
- поддержание стабильности содержаний восприятия в противовес помехам, создаваемым собственным движением;
- объединение интероцептивной информации о движениях тела и саморегулируемых потребностях тела (напр., температура, жажда, голод);
- поддержание высокоуровневого принятия решений с абстрагированием от нерелевантных сенсомоторных деталей их реализации.
Разбор нейроанатомических деталей в аргументации Меркера выходит за рамки темы настоящей статьи (Merker 2005). Он приходит к выводу, что системы, решающие вышеперечисленные проблемы — порождающие сознание — возникли у раннего предка позвоночных. Следовательно, сознание является одновременно и древним, и широко распространенным среди ныне живущих позвоночных.
6.6 Беспозвоночные
На фоне принципиальных различий анатомии и физиологии позвоночных и беспозвоночных возникают дополнительные сложные проблемы, связанные с вопросом наличия сознания за пределами родственной группы позвоночных.
В этом случае намного сложнее использовать стратегию выявления гомологически и функционально схожих структур и процессов (Seth et al. 2005), которая обосновывает уверенность в том, что сознание встречается и у других животных — ее разделяют такие исследователи, как Кабанак (Cabanac 2009), Сет и др. (Seth et al. 2005), Меркер (Merker 2005) и Панксепп (Panksepp 2005).
Родственная группа позвоночных представляет собой лишь один из примерно 34 известных типов — это древняя группа животных с различной базовой анатомической организацией и порождающими ее процессами развития. Каждый из этих типов происходит из относительно простой структуры (малочисленные виды тканей и минимальная центральная нервная система с ограниченными сенсорными возможностями).
Следовательно, такие беспозвоночные, как головоногие моллюски (напр., осьминоги и кальмары) и членистоногие (напр., ракообразные, насекомые и пауки) — существа, которые устроены достаточно сложно, чтобы привлечь внимание тех, кто интересуется сознанием животных, — развили свою сложность независимо от позвоночных, и даже независимо друг от друга.
Сегодня только три типа (позвоночные, членистоногие и моллюски) включают животных со сложными подвижными телами (см. Trestman 2013a), которые имеют следующие характеристики:
- развитые суставные конечности;
- контролируемые движения с большим количеством степеней свободы;
- периферийные органы чувств (напр., «настоящие» глаза);
- анатомическая способность к активной подвижности, управляемой периферийными органами чувств (плавники, ноги, движение с помощью реактивной струи и т.д.);
- анатомическая способность к активным манипуляциям объектами (напр., при помощи челюстей, руками, щупалец, ротовых частей с мелкомоторным управлением).
По Трестману, эволюция сложных подвижных тел требует способности к интегрированному, телесно воплощенному пространственному познанию, и эта способность формировалась независимо в каждом из трех типов, у которых она в настоящий момент присутствует (позвоночные, членистоногие и моллюски).
Такую линию рассуждения можно подкрепить, рассмотрев роль темпоральной интеграции перцептивной информации в сознании, а также в выборе действий и в объектно-ориентированном восприятии (Trestman 2013b). Возможно, сознание формировалось независимо у представителей каждой их этих трех родственных групп при их переходе от относительно простой морфологии тела (подобной той, которая характерна для червей) к сложным подвижным телам.
В контексте вопросов о сознании особое внимание было уделено одному из типов беспозвоночных животных, а именно двужаберным головоногим — осьминогам, кальмарам и каракатицам.
Эти животные имеют крупный мозг и известны своей хитростью, выдающимися способностями маскировки, а также гибкими стратегиями охоты. Матер утверждает, что головоногие демонстрируют множество поведенческих индикаторов сознания, включая комплексное обучение и пространственную память, а также явную способность к игре (Mather 2008). Как Меркер, так и Эдельман и др. утверждают, что можно привести убедительные предварительные доводы в пользу наличия сознания у головоногих — хотя они и подчеркивают те ограничения, которые накладывают на наше понимание имеющиеся различия между анатомией и физиологией головоногих и позвоночных (Merker 2005; Edelman et al., 2005, 2009).
Другой тип, которому уделяется особое внимание — это членистоногие, к которым относятся насекомые, ракообразные, пауки и многие другие, менее знакомые нам животные. Это древняя и чрезвычайно разнообразная группа животных, поэтому любые обобщения следует делать крайне осторожно. Членистоногие были среди самых первых животных, у которых сформировались сложные подвижные тела — и, соответственно, сформировался мозг, способный адаптивно управлять сложными адаптивными телами (Trestman 2013a).
Таким образом, если функция сознания заключается в решении проблем, возникающих при управлении сложными подвижными телами (ср. Merker 2005), то возможно оно сформировалось на ранних этапах эволюции членистоногих, у общего предка всех ныне живущих членистоногих.
Немногие исследования непосредственно стремятся дать ответ на вопрос о наличии сознания у членистоногих. Соответствующие эмпирические работы:
- Исследование Элвуда и Аппеля (Elwood & Appel 2009), которое показывает, что крабы-отшельники помнят негативное событие (удар током) и могут прибегать к воспоминанию при последующем принятии контекстно обусловленных решений. По всей видимости, это удовлетворяет определению того, что представляют собой «мотивированные уступки» в поведении, данному Кабанэк и Кабанэк (Cabanac & Cabanac 2009), и что с их точки зрения является индикатором сознания.
- Исследование Джексона и его коллег (рецензия в Jackson and Wilcox 1998), документирующее разнообразие впечатляющего поведения пауков-скакунов, в том числе:
- Обход препятствий и другие формы видимого планирования (признаны потенциальными индикаторами сознания в Cabanac et. al. 2009)
- гибкая, контекстуально обусловленная перестройка поведения от роли хищника к роли жертвы
- тактика обмана и ложных маневров
- Исследования, проводимые на пчелах, выявили распознавание образов, наличие понятий «одно и то же» и «другое», навигацию, коммуникацию, зрительную рабочую память (обзор в Srinivasan 2010), а также искажения, связанные с когнитивным состоянием и настроением (Mendl et al. 2011).
Еще одим вариант — сознание сформировалось еще раньше в ходе эволюции животных и в еще большей степени распространено среди животных, а стало быть, имеет еще более фундаментальную функцию в их жизни. Гинзбург и Яблонка характеризуют примитивную форму «суммарного ощущения» как побочный продукт, имеющийся даже у самых простых нервных сетей животных (Ginsburg & Jablonka 2007a,b). Они утверждают, что по мере приспособления этих состояний к обучению и мотивации, они приобретали функциональные свойства «базового сознания». Если это верно, то сознание могло возникнуть у членистоногих, моллюсков и позвоночных не независимым образом, а лишь однажды — у общего предка этих древних групп, на очень ранних этапах эволюции животных.
Исследования сознания животных: актуальные вопросы
На фоне постепенного ослабления бихевиористских ограничений в психологии и этологии, а также с появлением независимых успехов в неврологии, наблюдается значительное увеличение числа исследований животных, касающихся их сознания.
Некоторые из этих исследований сосредоточены на вполне определенных видах опыта (напр., боль), в то время как другие сосредоточены на когнитивных способностях (напр., осведомленность о себе), которые по всей видимости имеют прочную корреляцию с человеческим сознанием.
Данный раздел содержит краткие обзоры некоторых основных направлений этих исследований. Задача заключается в том, чтобы предоставить некоторые отправные точки для дальнейшего изучения научной литературы.
Боль и страдание животных
Неудивительно, что существует значительное количество (пусть и противоречивой) научной литературы, посвященной боли животных, учитывая то, что значение животной модели боли в клинических медицинских исследованиях, а также то, что большинство трактовок наших этических обязательств по отношению к животным отводят боли центральную роль (см. обзор: Mogil 2009). Недавние доклады Совета Наффилда по биоэтике в Великобритании (Nuffield Council 2005 [esp. ch. 4]) и Института исследований лабораторных животных Национальной академии наук США (ILAR 2009) охватывают проблематику определения боли, а также физиологических, неврологических и поведенческих свидетельств наличия боли у нечеловеческих животных. В них также проводится различие между болью с одной стороны и дистрессом, страданиями с другой (см. также Bermond 2001). Ранее единый доклад ILAR о распознавании и облегчении боли и дистресса был разделен на два отдельных доклада, хотя научное исследование дистресса пока находится, скорее, в зачаточном состоянии (тем не менее см. Dawkins 1985, Farah 2008).
Ноцицепция — чувствование болевых раздражителей — одна из самых примитивных сенсорных способностей. Нейроны, функционально специализирующиеся на ноцицепции, описаны у беспозвоночных, таких как медицинская пиявка и морская улитка Aplysia californica (Walters 1996).
Поскольку ноцицепторы встречаются у очень широкого круга видов и функционально активны даже у децеребральных животных и животных со спинальным трансектом, их наличие и функционирование у того или иного вида не дает практически никаких прямых доказательств наличия феноменально осознанного опыта боли. Теория контрольных ворот Мелзака и Уолла (Melzack and Wall 1965) описывает механизм, с помощью которого сигналы, идущие от мозга «сверху вниз», модулируют ноцицепцию «снизу вверх», что позволяет разграничить чувствуемую боль и ноцицепцию.
Смит и Бойд рассматривают свидетельства способности животных чувствовать боль с точки зрения того, связаны ли ноцицепторы с центральной нервной системой, имеются ли эндогенные опиоиды, влияют ли анальгетики на реакции, а также сходны ли последующие поведенческие реакции с теми, что наблюдаются у человека (Smith & Boyd 1991; ср. Varner 1998 [p. 53, table 2.3 — более новая версия представленной у Смита и Бойда]).
Основываясь на этих критериях, Варнер присоединяется к Смиту и Бойду и предварительно заключает, что наиболее очевидная граница между организмами, осознающими боль, и организмами, не способными сознательно испытывать боль — это граница между позвоночными и беспозвоночными. Тем не менее Элвуд и Аппель провели эксперимент на крабах-отшельниках и считают, что его данные свидетельствуют о том, что эти ракообразные испытывают боль и помнят этот опыт (Elwood & Appel 2009). Варнер, скорее, сомневался относительно свидетельств в пользу наличия сознательной боли у «нижних» позвоночных: рыб, рептилий и земноводных. Однако Аллен утверждает, что последовавшие исследования демонстрируют: открытия направлены на выявление бόльшего количества сходств между разнообразными видами, относящимися к различным таксономическим классам, особенно в области анатомии и физиологии ноцицептивной и болевой систем (Allen 2004).
Общепризнано, что болевая система млекопитающих имеет как сенсорные, так и аффективные каналы, и что они могут быть до некоторой степени разъединены –как фармакологически, например, при помощи морфина, так и при помощи хирургического вмешательства. В этом отношении передняя поясная кора (ППК) является особенно важной структурой мозга млекопитающих (Price 2000). Аллен и др., а также Шрайвер утверждают, что эта описанное разъединение дает возможность эмпирически оценить аффективный компонент сознания животных (Allen et al. 2005, Shriver 2006), а Фарах использует ее для того, чтобы различать страдание и «просто боль» (Farah 2008).
Тем не менее детальный анализ других таксономических групп может выявить важные анатомические различия. Роуз утверждает, что поскольку у рыб передняя поясная кора (ППК) отсутствует, боль может их не беспокоить (Rose 2002).
Это противоречит позиции Снеддона и др., которые утверждают, что существуют релевантные поведенческие и физиологические свидетельства, позволяющие приписывать боль рыбам (Sneddon et al. 2003). ППК важна для млекопитающих, однако сохраняется вероятность того, что другие таксоны могут иметь функционально схожие структуры, такие как corticoidea dorsolateralis у птиц (Atoji & Wild 2005; Dubbeldam 2009). Животные с выключенными генами также дают дополнительные сведения об аффективных аспектах боли (см. Shriver 2009 — обзор и применение этих результатов для благополучия животных).
Наконец, следует отметить, что за последнее десятилетие произошел серьезный сдвиг в ветеринарной практике, касающийся отношения к боли животных.
Некоторые даже утверждают, что животные обладают нейробиологическими механизмами, ответственными за фантомные боли в конечностях и невропатические боли (боль при отсутствии явных повреждений тканей или заболевания), потому такие состояния можно обнаружить у нечеловеческих животных — и вылечить их (Mathews 2008).
Эмоции животных
Конечно, представление о наличии эмоций у животных занимает видное место в творчестве Дарвина, особенно в его книге «О выражении эмоций у человека и животных» (2001 [1872]). Стремление рассматривать животных как эмоциональных существ (а человека, напротив, — как существо, наделенное рациональностью, способное преодолевать эмоции) прослеживается по меньшей мере до древнегреческой философии.
Конрад Лоренц, похоже, придерживался сходных взглядов, вторя утверждению Оскара Хайнрота, что «животные — очень эмоциональные люди с очень ограниченным интеллектом» (Lorenz 1971b: 334). Сегодня более модно рассматривать эмоции как важный компонент интеллекта. Вне зависимости от достоинств этой точки зрения, научное изучение эмоций животных приобретает все большие масштабы. В начале ХХ века физиологи признали значимость эмоций для поведения животных, пускай тогда в их сообществе и не велись споры о наличии у животных сознания.
По мнению Дрора, хотя физиологи были вынуждены заниматься проблемой эмоциональных искажений, попытки рассматривать эмоции в качестве самостоятельного предмета исследования так и не смогли оформиться институционально, в виде журнала или как-либо еще (Dror 1999: 219).
Относительно недавно Яак Панксепп (Panksepp 2004, 2005) стал осуществлять исследовательскую программу, которую он называет «аффективной неврологией» и которая включает непосредственное исследование эмоций животных (Panksepp 2004).
Один из примеров — экспериментальное исследование крыс, которые «смеются» и ищут дальнейшего контакта в ответ на щекотание людей (Panksepp & Burgdorf 2003). За несколько десятилетий его исследование (см. обзор в Panksepp 2005) смогло пролить свет на нейро- и молекулярно-физиологические основы нескольких «базовых эмоциональных систем», включая «поиск», «страх», «ярость», «похоть», «забота», «игра», «паника». Панксепп утверждает, что они разделяются всеми млекопитающими и могут встречаться среди более широкого круга позвоночных животных.
Суфка и соавторы предположили, что модельные животные, имеющие нейропсихиатрические расстройства, могут также способствовать экспериментальному исследованию эмоций у животных (Sufka et al. 2009).
Со стороны ученых как Сматс, так и Бекофф отстаивают приписывание сознательных эмоций животным, хотя на основе скорее анекдотического, нежели экспериментального подхода (Smuts 2001, Bekoff 2007). Бекофф придает большое значение игровому поведению как показателю наличия эмоций у животных (эту идею подхватили и в Cabanac et al. 2009).
Еще одна тема текущих исследований — это эмпатия у животных (напр., Preston & de Waal 2002). Лэнгфорд и др. высказываются в пользу наличия эмпатии у мышей на основе проведенных экспериментов, в которых мыши, наблюдавшие за соседом по клетке, получавшим болевой раздражитель или испытывавшим боль, оказываются более чувствительны к болезненным раздражителям, чем контрольные мыши, наблюдавшие за незнакомой мышью в такой же ситуации (Langford et al. 2002). Бирн и др. высказываются в пользу наличия эмпатии у слонов, поскольку такой вывод лучше всего объясняет некоторые из их способностей — такие как распознавание одушевленности, целеустремленность, а также оценка тех физических способностей и эмоциональных состояний у других слонов, которые отличаются от их собственных (Byrne et al. 2008).
Следует отметить, что почти все исследования, посвященные эмоциям животных — как теоретические, так и эмпирические — ограничиваются млекопитающими или хотя бы амниотами (см. примечательное недавнее исключение: Mendl et al. 2011). Эти исследования разрабатывают определенные глубинные гомологии, исследуя структуру мозга и молекулярную нейрофизиологию (с акцентом на гормоны и нейромедиаторы) и выдвигают доводы в пользу того, что животные в указанных таксономических группах имеют те же эмоции, что и люди (как и людям, им присуще поведение, демонстрирующее эмоции, связанные в физиологической основе с определенными закономерностями).
Такой подход неприменим к тем животным, которые имеют очень отдаленную связь с нами — к беспозвоночным. Предки позвоночных отделились от остальных животных типов в то время, когда все животные были еще относительно просты с точки зрения структуры, типов тканей, количества нейронов, способностей тела к перемещению и иных форм поведения. Формирование сложных физиологических систем происходило в различных типах независимо. Потому физиологические системы, лежащие в основе эмоций у представителей других типов, могут быть очень разными (если вообще существуют), а следовательно, их трудно идентифицировать в рамках и непосредственных физиологических наблюдений, и наблюдения за поведенческими проявлениями эмоций.
Возможно также, что набор эмоций у представителей других типов отличается, что еще больше затрудняет задачу выявления эмоций у беспозвоночных. (Дальнейшее обсуждение и дополнительные ссылки на научные исследования эмоций и эмпатии у животных можно найти в разделе об эмоциях и эмпатии в статье о познании животных.)
7.3 Феноменология восприятия
Представление о том, что скрупулезные психофизические исследования могли бы помочь нам понять природу субъективного опыта мира, имеющегося у исследуемых, прослеживается по меньшей мере до экспериментальных тестов Дональда Гриффина, посвященных ограничениям способности эхолокации у летучих мышей. Оно также лежит в основе другого представления — будто бы знание о том, что угол обзора лошади составляет 360о или что у хищников на сетчатке каждого глаза по два углубления, может рассказать нам нечто о качествах опыта животных (о том, как им представляется мир — если мы считаем, что такой опыт у них есть).
Исследование нейронов добавляет к научному пониманию природы восприятия еще один пласт анализа. Так, Леопольд и Логотетис использовали данные о нейронах для обоснования выводов о том, что воспринимают обезьяны в условиях бинокулярного соперничества (Leopold & Logothetis 1996; см. также Myserson et al. 1981, Rees et al. 2002). Для Леопольда и др. учет нейронных данных может использоваться для подтверждения того, что обезьяны, получившие противоречащие визуальные стимулы, дают невербальные «сообщения» (Leopold et al. 2003).
Феномен слепозрения — ложной слепоты, формы бессознательного зрительного восприятия, которое возникает при повреждении определенных участков первичной зрительной коры, — был также исследован у обезьян, подвергшихся хирургическому вмешательству (Stoerig & Cowey 1997). Исследования показали близкое сходство между теми поражениями, которые имелись у обезьян и у человека: между теми участками поля зрения, которые могут быть обработаны только бессознательно, и теми, в отношении которых исследуемые остаются сознательными. Невербальный подход к оценке зрительной осведомленности был в дальнейшем подтвержден Стоеригом и др. (Stoerig et al. 2002).
Испытуемые со слепозрением — как люди, так и обезьяны — не реагируют спонтанно на предметы, представленные в их скотоме (участке поля зрения, о существовании которого они визуально не осведомлены), но они должны быть обучены реагировать при помощи «вынужденной реакции» (Stoerig & Cowey 1997).
Большинство примеров делает упор на визуальное восприятие, что несомненно отражает ангажированность в пользу приматов. Мы, человеческие приматы, в высокой степени визуальные существа, и, как утверждал Нагель (2003), мы сталкиваемся со значительными препятствиями, когда пытаемся представить (опять же, зрительная метафора) субъективный опыт других существ в тех модальностях, в которых человек слаб или полностью неодарен.
Существуют хорошие исследования нейроэтологов, рассматривающие психофизику эхолокации у летучих мышей и дельфинов или сенсорные способности электрических рыб, однако вопросы субъективной осведомленности в этой литературе, как правило, не затрагиваются.
Скрупулезный анализ нейронных коррелятов сознания может выявить тонкие различия между животными разных видов, как это было в случае изучения боли у животных. Так, сообщается, что поведение макака-резуса весьма сходно с поведением человека в условиях бинокулярного соперничества, однако
…активность большинства нейронов в зрительной зоне V1 и рядом с ней отражает в первую очередь свойства стимула, а не сознательное восприятие объекта, о котором сообщает животное… Тем не менее несколько исследований нейровизуализации у людей представили свидетельства в пользу значительной роли зрительной зоны V1 в условиях бинокулярного соперничества и, косвенным образом, в пользу зрительной осведомленности. (Rees et al. 2002)
Примечательно тем не менее, что с их точки зрения поведенческие свидетельства могут безо всяких проблем трактоваться как признак того, что у обезьян имеется сознательное восприятие.
Ментальное путешествие во времени
В литературе развернулась дискуссия о том, способны ли другие животные, подобно людям, думать о прошлых и будущих событиях. Суддендорф и Корбаллис (Suddendorf and Corballis 1997, 2007) утверждают, что «ментальное путешествие во времени» присуще исключительно человеку и действительно играет важную роль в объяснении того, что является когнитивно уникальным для него, включая способность к языку и культуре. Многие млекопитающие, птицы и рыбы демонстрируют такие формы поведения, которые, похоже, предполагают прогнозирование — заготовка пищи, создание гнезд, использование инструментов, миграция. Так, тайры — представители семейства куньих, обитающие в Центральной и Южной Америке, — прячут ветки бананов внутри бромелий и достают их только после того, как бананы созреют (Soley et al. 2011).
Скептики сразу же указывают на то, что многие из примеров показывают вовсе не поведение, опосредованное «когнитивными» процессами (эпизодической памятью, проницательностью или пониманием). Они демонстрируют сугубо (а) «инстинктивные» паттерны фиксированного действия, сформированные в процессе естественного отбора, либо же (б) результат классического или оперантного обучения. Однако свидетельства о наличии новизны, гибкости и способности приспосабливаться в зависимости от ситуации часто ставят критику этой гипотезы под сомнение. Например, тайры делают запасы бананов нескольких видов и точно оценивают, достаточно ли созрел банан такого-то вида или ему необходимо еще дозреть. Это работает и в отношении домашних бананов, с которыми тайры не имели контакта в ходе своей эволюции (Soley et al. 2011).
Разнообразные и тщательные экспериментальные исследования демонстрируют впечатляющие способности к интегрированной памяти «что-где-когда» — животные вспоминают детали события вместе с его локацией и временем. Такое исследование было впервые проведено Клейтоном и коллегами на хохлатых сойках, акцент делался на поведении по запасанию пищи, когда птицы закапывают пищу, а затем извлекают ее.
Например, если хохлатым сойкам довольно долгое время не дают извлечь свои запасы, то они откопают в итоге только нескоропортящиеся запасы (в исследовании — арахис), и проигнорируют запасы той пищи, которая обычно является более предпочтительной, но скоропортящейся (в исследовании — мучные черви; см. Clayton et al. 2003). Недавние исследования также документально подтверждают наличие «эпизодоподобной памяти» у крыс (Crystal 2009), а также явную способность планировать будущее (даже в новых условиях, что нельзя оправданно сбросить со счетов как «простой инстинкт»), имеющуюся у некоторых животных, в том числе и у нечеловеческих приматов, птиц, крыс и прочих млекопитающих (см. Feeney et al. 2011 как пример недавнего экспериментального исследования; см. обзор и обсуждение: Roberts 2012).
Этот спор несколько усложняется в связи с тем, что сторонники приписывания ментального путешествия во времени исключительно человеку полагаются на такие его описания, которые нагружены феноменологическими, интроспективными или интуитивными описаниями работы их собственной психики (напр., Tulving 1985; Suddendorf and Corbalis 2007). Однако исследователи поведения животных для подтверждения своих утверждений должны полагаться на строгие стандарты поведенческих свидетельств.
Исследователи поведения животных, как правило, осмотрительны в своих интерпретациях. Они ограничивают свои суждения операционализируемыми терминами (память «что-где-когда», «эпизодоподобная» память) и не делают заявлений о природе опыта, который может быть задействован животным при выполнение той или иной задачи.
Такой пример Бакнер (Buckner 2013) назвал «антропофабуляцией». Вопрос о человеческой осведомленности о времени — в какой степени и каким образом оно отличается от осведомленности у других животных — остается открытым и актуальным для текущих исследований.
Самосознание и метакогниции
Истоки систематического изучения самосознания и теории психики у нечеловеческих животных обнаруживаются в том подходе к изучению самосознания, который был впервые предложен Гэллапом (Gallup 1970). Как Гэллап обосновал связь узнавания себя в зеркале с осведомленностью о себе, обсуждалось выше. Идею эксперимента подсказало наблюдение, хорошо известное психологам-компаративистам — что шимпанзе (после периода приспособления) используют зеркала для изучения своего собственного образа.
Гэллап использовал эти наблюдения и разработал широко тиражируемый протокол, позволяющий научно определить, является ли объектом интереса животного только зеркальное изображение как таковое или же изображение служит объектом интереса, выступая проводником к самому животному. Гэллап взял тех шимпанзе, которые были уже хорошо знакомы с зеркалами, анестезировал их и нанес на лоб метки, а в контрольной группе только анестезировал их.
После пробуждения животные с меткой, которым было разрешено видеть себя в зеркале, дотрагивались до своего лба в районе метки значительно чаще, чем те подопытные, которые были без метки или которым не было разрешено смотреть в зеркало.
Обычно сообщается, что шимпанзе всякий раз «проходят» тест с зеркалом и метками, но обзор научной литературы (Shumaker & Swartz 2002) показывает, что из 163 протестированных шимпанзе только 73 демонстрировали поведение, когда они трогали метку (хотя среди этих животных были значительные различия в возрасте и в опыте взаимодействия с зеркалами). Авторы исследования также сообщают, что такое поведение демонстрировали 5 из 6 протестированных орангутангов и 6 из 23 горилл. Предполагается, что редкость поведения с касанием метки у горилл связана с тем, что в экспериментах не было социально-значимого прямого зрительного контакта.
Очень мало свидетельств узнавания себя в зеркале в случае тех приматов, которые не относятся к человеческому роду или к высшим приматам. Сам Гэллап считал, что это явление присуще исключительно высшим приматам. Он одним из первых выступил с критикой доклада Хаузера, в котором говорилось, что эдиповы тамарины благодаря зеркалу вовлекаются в самонаправленное поведение, если предварительно окрасить их характерные белые хохолки в неоновые цвета — такое воздействие предположительно более заметно, чем использованная Гэллапом красная метка, по утверждению Хаузера и соавторов (Hauser et al. 1995).
Столкнувшись с критикой Гэллапа, Хаузер не смог воспроизвести свои первоначальные результаты (Hauser et al. 2001). Тем не менее мысль о том, что протокол Гэллапа использует не слишком заметный для обезьян стимул, продолжает иметь некоторый вес. Например, Раджала и др. представили количественные и видеографические свидетельства того, что макаки-резусы с имплантатами в голове используют зеркала для осмотра имплантатов, а также других частей тела, которые они обычно не видят (Rajala et al. 2010).
Модифицированные версии эксперимента Гэллапа также проводились с представителями видов, не являющихся приматами. Известно, что Эпштейн и др. тренировали голубей таким образом, чтобы они клевали метку на собственном теле, которая была видна только в зеркале (Epstein et al. 1981). Они использовали данные этого эксперимента, чтобы поставить под сомнение приписывание «осведомленности» на основе теста с зеркалом и меткой, и сами при объяснении результатов ссылались на ассоциативное обучение. Гэллап и др. отвергают заявленную эквивалентность экспериментов и отмечают, что шимпанзе не были обучены прикасаться к меткам до проведения теста (Gallup et al. 2002). Райс и Марино представили свидетельства наличия зеркального самоузнавания у афалин (Reiss & Marino 2001).
Они модифицировали протокол Гэллапа (не использовали анестезию) и сделали вывод о наличии самоузнавания на основе скручиваний тела перед зеркалом (поскольку самокасание в случае дельфинов анатомически невозможно). Этот тезис был оспорен (напр., Wynne 2004). Тест с зеркалом и меткой продолжает оставаться областью активных исследований различных видов, включая слонов (Plotnik et al. 2006) и сорок (Prior et al. 2008). Многие комментаторы отмечали, что зеркальный тест, возможно, не вполне справедлив для тех видов, которые в бόльшей степени полагаются не на зрение, а на ощущения (Mitchell 2002; Bekoff and Burghardt 2002).
Любопытное направление исследования того, что животные знают о собственных ментальных состояниях, рассматривает их поведение в ситуациях когнитивной неопределенности. Когда приматам и дельфинам дается «спасительный» ответ, позволяющий им избежать сложного выбора, они выбирают вариант спасения, что очень похоже на человека (Smith et al. 2003). Животные, у которых нет варианта спасения и которые вынуждены реагировать на сложные сравнения, справляются хуже, чем те, у которых есть выбор и которые сами отказались от варианта спасения. Этот факт использовали в качестве аргумента в пользу того, что существует некий вид высокоуровневого понимания себя. Философы критиковали первоначальные эксперименты из-за интерпретации второго уровня (напр., Carruthers 2008), психологи — из-за методологии (см. обзор: Crystal & Foote 2009). Впрочем, другие попытки установить наличие метакогниций у животных, не обладающих языком, могут избежать подобной критики (Terrace & Son 2009).
В литературе о когнитивных способностях человека «метакогницией» называют осведомленность о том, что человек знает, и она ассоциируется с «чувством знания».
По их мнению, достигнутые ими результаты уже показывают, что «животные имеют функциональные особенности такие же как или сходные с сознательным познанием человека» (Smith et al. 2003; Smith 2009). Вдобавок они ставят вопрос о том, что это может нам рассказать о феноменальных особенностях познания. По Брауну, исследования дельфинов не дают свидетельств в пользу наличия у них теории психики, но говорят о наличии у них сознания (Browne 2004), особенно если использовать трактовку Ликана, описанную выше. Понятие метакогниции может оказаться актуальным в случае вопросов о сознании доступа. (Дополнительное обсуждение осведомленности о себе и метакогниций см. в разделе, посвященном теории психики и метакогнициям в статье о познавательной способности животных.)
Итоги
Статья вроде нашей, скорее, поднимает вопросы, нежели дает ответы — однако в противном случае сама тема едва ли представляла бы интерес с философской точки зрения.
Очевидно, что для многих философов вопрос о наличии сознания у животных не является второстепенным. Растет интерес к сознанию животных в самых разных направлениях философии, в том числе в этике, философии сознания, а также в философии науки. Философы, работающие в этих областях, уделяют все больше внимания отдельным тонкостям научной теории, ее методам и результатам.
Впрочем, с обеих сторон остаются и критики: по одну сторону находятся те, кто по-прежнему считает, будто субъективные феномены лежат за рамками научного исследования, по другую — те, кто полагает, будто наука и философия еще недостаточно продвинулись на пути признания того, что животные обладают сознанием (или же продвигаются слишком медленно). Набор аргументов у каждой из сторон не близится к истощению.
Примечания автора
1. См. разбор филогенетического распределения механизмов, отвечающих за боль: Varner 1999: 51–54. Однако стоит быть внимательным в отношении таблицы на с. 53, в соответствии с которой специальные нейроны ноцицепторы имеются только у позвоночных. Все же существование ноцицепторов у морских моллюсков надежно установлено (Walters, E. T., [1992] “Possible clues about the evolution of hyperalgesia from mechanisms of nociceptive sensitization in aplysia.” Hyperalgesia and Allodynia (ed. W. D. Willis, Jr.), N.Y.: Raven Press).
2. Выражение «испытывать боль» для большинства читателей означает сознательный опыт, однако не все с этим соглашаются. Например, Каррутерс полагает, что хотя животные действительно испытывают боль, они не испытывают ее сознательно (Carruthers 1998b). Иными словами, в его несколько парадоксальной формулировке отрицается, что это чувство чувствуется каким бы то ни было образом. Тем не менее многие философы возразят на это, что само понятие неосознанной боли является противоречивым, поскольку боль по своей сути осознанна.
3. Поскольку наличие чувствительности у животных важно для этики, некоторые наиболее подробно изложенные формулировки эволюционно подкрепленного аргумента сходства можно обнаружить в литературе, посвященной защите прав животных и их благополучия (напр., Singer 1975/1990; Regan 1983; Andrews 1996; Varner 1999).
4. Интересно отметить — было установлено, что у многих видов, особенно среди китообразных и перелетных птиц, полушария мозга погружаются в сон попеременно. Такая адаптация позволяет морским млекопитающим не утонуть, а птицам – продолжать полет. Нам неизвестно ни одно систематическое исследование, посвященное работе перцептивных и когнитивных способностей в условиях, когда половина мозга находится в состоянии сна.
5. У этой проблемы много формулировок: Аллен и Бекофф называют ее «проблемой психики других видов» (the other species of mind problem, Allen & Bekoff 1997), а Принц — «проблемой субъекта» (The Who Problem, Prinz 2005).
6. Однако свидетельства в пользу этого не вполне однозначные. Напр., Гювен Гюзельдере (Güzeldere 1995) сообщает о переписке с Дэвидом Армстронгом, в которой тот пишет следующее:
[Güzeldere, G., (1995), “Is consciousness the perception of what passes in one's own mind?” Conscious Experience (ed. Th. Metzinger), Paderborn: Schöningh/Imprint Academic, 1995, fn. 22.]
Ликан в своей статье для журнала Psyche, где были опубликованы материалы симпозиума по сознанию животных, комментируя позицию Каррутерса (Carruthers 1998a), пишет следующее:
Так высказываются теоретики, разделяющие теорию высокоуровневого опыта, и едва ли это горячее одобрение наличия сознания у животных.
7. Хаузер, Хомский и Фитч (Hauser, Chomsky, and Fitch 2002) утверждают, что исследования в рамках сравнительного языкознания столкнулись с затруднениями при попытке выделить три различных направления сравнения человеческой способности к языку в широком смысле (СЯШ) и других форм животной коммуникации. Эти три направления они обозначили как «сенсомоторное», «концептуально-интенциональное» и «СЯУ-рекурсия» (СЯУ — способность к языку в узком смысле). Их статья содержит полезный обзор данных, свидетельствующих о значительной эволюционной гомологии и гомоплазии в первых двух направлениях.
Хаузер и Фитч (Hauser & Fitch 2004) провели дополнительное исследование третьего направления и обнаружили следующее: тамарины способны различать последовательности, соответствующие грамматике с конечным числом состояний, которая допускает произвольные повторения простого паттерна (напр., ABn), но они не могли справиться с фразово-структурной грамматикой (напр., AnBn).
Однако выяснилось, что скворцовые могут различать паттерны AnBn (Gentner et al. 2006; хотя эти авторы, возможно, переоценили полученные результаты, посчитав их примером освоения рекурсивного правила, раз фразово-структурные грамматики не требуют полной рекурсии). Другие возможные эволюционные соотношения человеческого языка и коммуникации животных обсуждаются также философами: Allen and Saidel 1998, Trout 2001; лингвистическая перспектива: Anderson 2004.
8. Этот аргумент очень напоминает аргумент Дэвидсона (Davidson 1975), который опровергал наличие мышления у животных на основе того, что способности мышления должно предшествовать понятие мышления. Нам неизвестно, возникал ли у него соблазн обратить этот аргумент против наличия у животных чувствительности.
Библиография
· Дарвин, Ч. (1874). Происхождение человека и подбор по отношению к полу, пер. И. М. Сеченова, С.-Петербург.
· Дарвин, Ч. (2001). О выражении эмоций у человека и животных. Санкт-Петербург,.
· Лавджой, А. (2001). Великая цепь бытия, Москва.
· Нагель, Т. (2003). «Каково быть летучей мышью?» Хофштадтер, Д. & Деннет, Д. (ред.) Глаз разума: Фантазии и размышления о самосознании и о душе, Самара, с. 349–360.
· Юм, Д. (1996). «Трактат о человеческой природе». Соч.: В 2 т., Москва, т. 1.
· Akins, K. A. (1993). A Bat Without Qualities. In M. Davies & G. Humphreys (eds.), Consciousness (pp. 258–273). Oxford: Blackwell.
· Alcock, J. (1992). Review of Griffin 1992. Natural History, 101(9), 62–65.
· Allen, C. (1992a). Mental Content. British Journal for the Philosophy of Science, 43(4), 537–553.
· ––– (1992b). Mental Content and Evolutionary Explanation. Biology and Philosophy, 7(1), 1–12.
· ––– (2004). Animal Pain. Noûs, 38(4), 617–643.
· ––– (2013). Fish cognition and consciousness. Journal of Agricultural and Environmental Ethics, 26(1), 25–39.
· Allen, C., & Bekoff, M. (1997). Species of Mind: The Philosophy and Biology of Cognitive Ethology. Cambridge, MA: MIT Press.
· Allen, C., Fuchs, P. N., Shriver, A. & Wilson, H. (2005). Deciphering Animal Pain. In M. Aydede (ed.), Pain: New Essays on the Nature of Pain and the Methodology of its Study (pp. 352–366). Cambridge, MA: MIT Press.
· Allen, C., & Grau, J., & Meagher, M. (2009). The Lower Bounds of Cognition: What Do Spinal Cords Reveal? In J. Bickle (ed.), The Oxford Handbook of Philosophy of Neuroscience (pp. 129–142). Oxford: Oxford University Press.
· Anderson, S. R. (2004). Doctor Dolittle's Delusion. New Haven: Yale University Press.
· Andrews, K. (1996). The first step in the case for great ape equality: the argument for other minds. Etica & Animali (Special issue devoted to The Great Ape Project), 8, 131–141.
· ––– (2012). Do Apes Read Minds? Toward a New Folk Psychology. Cambridge, MA: MIT Press.
· Ârhem, P., & Liljenström, H., & Lindahl, B. I. B. (2002). Evolution of Consciousness: Report of Agora Workshop in Sigtuna, Sweden, August 2001. Journal of Consciousness Studies, 9, 81–84.
· Armstrong, D. A. (1980). The Nature of Mind and Other Essays. Ithaca, NY: Cornell University Press.
· Atoji, Y., & Wild, J. M. (2005). Afferent and efferent connections of the dorsolateral corticoid area in comparison with connections of the temporoparieto-occipital area in the pigeon (Columbia livia). Journal of Comparative Neurology, 485, 165–182.
· Baars, B. J. (1997). In the Theatre of Consciousness: Global Workspace Theory, A Rigorous Scientific Theory of Consciousness. Journal of Consciousness Studies, 4, 292–309.
· Barrington, D. (1773). Experiments and observations on the singing of birds. Philosophical Transactions of the Royal Society, 63, 249–91.
· Bekoff, M. (2007). The Emotional Lives of Animals. Novato, CA: New World Library.
· Bekoff, M., & Allen, C. (1997). Cognitive Ethology: Slayers, Skeptics, and Proponents. In R. Mitchell, N. Thompson & H. Miles (eds.), Anthropomorphism, Anecdotes, and Animals (pp. 313–334). Albany, NY: State University of New York Press.
· Bekoff, M., & Allen, C., & Burghardt, G. M. (eds.). (2002). The Cognitive Animal.Cambridge, MA: MIT Press.
· Bermond, B. (2001). A neuropsychological and evolutionary approach to animal consciousness and animal suffering. Animal Welfare Supplement, 10, 47–62.
· Bertoloni Meli, D. (2013). Early Modern Experimentation on Live Animals. Journal of the History of Biology, 46(2), 199–226.
· Block, N. (1995). On A Confusion About a Function of Consciousness. Behavioral and Brain Sciences, 18, 227–47.
· Block, N. (2005). Two Neural Correlates of Consciousness. Trends in Cognitive Sciences, 9, 41–89.
· ––– (2011). Perceptual consciousness overflows cognitive access. Trends in cognitive sciences, 15(12), 567–575.
· Blumberg, M. S., & Wasserman, E. A. (1995). Animal Mind and the Argument from Design. American Psychologist, 50(3), 133–144.
· Boly, M., Seth, A. K., Wilke, M., Ingmundson, P., Baars, B., Laureys, S., Edelman, D., Tsuchiya, N. (2013). Consciousness in humans and non-human animals: Recent advances and future directions. Frontiers in Psychology, 4 (625), available online.
· Braithwaite, V. (2010). Do Fish Feel Pain? Oxford: Oxford University Press.
· Brigandt, I. (2005). The instinct concept of the early Konrad Lorenz. Journal of the History of Biology, 38(3), 571–608.
· Brown, C. (2015). Fish intelligence, sentience and ethics. Animal Cognition, 18, 1–17. doi:10.1007/s10071-014-0761-0
· Browne, D. (2004). Do dolphins know their own minds? Biology & Philosophy, 19, 633–653.
· Buckner, C. (2013). Morgan's Canon, meet Hume's Dictum: avoiding anthropofabulation in cross-species comparisons. Biology & Philosophy, 28(5), 853–871.
· Burghardt, G. (1985). Animal awareness: Current perceptions and historical perspective. American Psychologist, 40(8), 905–19.
· Burkhardt, R. W. Jr. (1997). The founders of ethology and the problem of animal subjective experience. In M. Dol, et al., Animal Consciousness and Animal Ethics: Perspectives from the Netherlands (pp. 1–13). Assen, the Netherlands: van Gorcum.
· ––– (2005). Patterns of Behavior: Konrad Lorenz, Niko Tinbergen and the Founding of Ethology. Chicago: University of Chicago Press.
· Byrne, R. W., et al. (2008). Do Elephants Show Empathy? Journal of Consciousness Studies, 15(10–11), 204–225.
· Byrne, R. W., & Whiten, A. (1988). Machiavellian Intelligence: social expertise and the evolution of intellect in monkeys, apes and humans. Oxford: Oxford University Press.
· Cabanac, A., & Cabanac, M. (2000). Heart rate response to gentle handling of frog and lizard. Behavioural Processes, 52, 89–95.
· ––– (2004). No emotional fever in toads. Journal of Thermal Biology, 29, 669–73.
· Cabanac, M., & Bernieri, C. (2000). Behavioral rise in body temperature and tachycardia by handling of a turtle (Clemys insculpta).. Behavioural Processes, 49, 61–68.
· Cabanac, M., & Cabanac, J., & Paren, A. (2009). The emergence of consciousness in phylogeny. Behavioural Brain Research, 2(198), 267–272.
· Cabanac, M., & Gosselin, F. (1993). Emotional fever in the lizard Callopistes maculates. Animal Behavior, 46, 200–202.
· Cabanac, A. J., & Guillemette, M. (2001). Temperature and heart rate as stress indicators of handled common eider. Physiology and Behavior, 74, 475–9.
· Carruthers, P. (1989). Brute experience. The Journal of Philosophy, 86(5), 258–269.
· ––– (1992). The Animals Issue. Cambridge: Cambridge University Press.
· ––– (1996). Language, Thought and Consciousness. Cambridge: Cambridge University Press.
· ––– (1998). Animal Subjectivity. Psyche, 4(3), 2377.
· ––– (1998). Natural Theories of Consciousness. European Journal of Philosophy, 6, 203–222.
· ––– (1999). Sympathy and subjectivity. Australasian Journal of Philosophy, 77(4), 465–482.
· ––– (2000). Phenomenal Consciousness: A naturalistic theory. Cambridge: Cambridge University Press.
· ––– (2004). Suffering without Subjectivity. Philosophical Studies, 121(2), 99–125.
· ––– (2008). Meta-cognition in Animals: A Skeptical Look. Mind & Language, 23, 58–89.
· Cavalieri, P., & Singer, P. (eds.). (1994). The great ape project: Equality beyond humanity.New York: St. Martin's Press.
· Chalmers, D. (2015). Panpsychism and panprotopsychism. In T. Alter & Y. Nagasawa (eds.), Consciousness in the Physical World: Perspectives on Russellian Monism. New York: Oxford University Press, pp 246-276.
· Chandroo, K. P., & Yue, S., & Moccia, R. D. (2004). An evaluation of current perspectives on consciousness and pain in fishes. Fish and Fisheries, 5, 281–295.
· Cheney, D. L., & Seyfarth, R. M. (1990). How Monkeys See the World: Inside the Mind of Another Species. Chicago: University of Chicago Press.
· ––– (2007). Baboon Metaphysics: The Evolution of a Social Mind. Chicago: University of Chicago Press.
· Clark, R. E., & Squire, L. R. (1998). Classical Conditioning and Brain Systems: The Role of Awareness. Science, 280, 77–81.
· Clayton, N. S., & Bussey, T., & Dickinson, A. (2003). Can Animals Recall the Past and Plan for the Future? Nature Reviews: Neuroscience, 4, 685–91.
· Clayton, N. S., & Yu, K. S., & Dickinson, A. (2001). Scrub Jays (Aphelocoma coerulescens) form integrated memories of the multiple features of caching episodes. Journal of Experimental Psychology: Animal Behavior Processes, 1, 17–29.
· Crist, E. (2002). The inner life of earthworms: Darwin's argument and its implications. In M. Bekoff, C. Allen & G. Burghardt (eds.), The Cognitive Animal (pp. 3–8). Cambridge, MA: MIT Press.
· Crystal, J. D. (2009). Elements of episodic-like memory in animal models. Behavioural Processes, 80(3), 269–277.
· Crystal, J. D., & Foote, A. L. (2009). Metacognition in animals. Comparative Cognition & Behavior Reviews, 4, 1–16.
· Damasio, A. (1999). Empathy and direct social perception. Review of Philosophy and Psychology, 3, 541–558.
· ––– (1999). The Feeling of What Happens. New York: Harcourt Brace.
· Darwin, C. (1881). The formation of vegetable mould, through the action of worms, with observations on their habits. London: John Murray.
· Davidson, D. (1975). Thought and Talk. In S. Guttenplan (ed.), Mind and Language (pp. 7–23). Oxford: Oxford University Press.
· Dawkins, M. S. (1985). The scientific basis for assessing suffering in animals. In P. Singer (ed.), In Defense of Animals (pp. 27–50). New York: Blackwell.
· Dawkins, M. S. (1993). Through Our Eyes Only? The Search for Animal Consciousness. New York: W. H. Freeman.
· ––– (2012). Why Animals Matter: Animal Consciousness, Animal Welfare, and Human Well-being. New York. Oxford: Oxford University Press.
· DeGrazia, D. (1997). Great apes, dolphins, and the concept of personhood. The Southern journal of philosophy, 3, 301–320.
· Dennett, D. C. (1969). Content and Consciousness. London: Routledge and Kegan Paul.
· ––– (1983). Intentional systems in cognitive ethology: The ‘Panglossian paradigm’ defended. Behavioral and Brain Sciences, 6, 343–390.
· ––– (1987). The Intentional Stance. Cambridge, MA: MIT Press.
· ––– (1995). Animal consciousness and why it matters. Social Research, 62, 691–710.
· ––– (1997). Kinds of Minds: Towards an Understanding of Consciousness. New York: Basic Books.
· Dretske, F. (1995). Naturalizing the Mind. Cambridge, MA: MIT Press.
· Dubbeldam, J. (2009). The Trigeminal System in Birds and Nociception. Central Nervous System Agents in Medicinal Chemistry, 9, 150–158.
· Edelman, D. B., & Baars, B. J., & Seth, A. K. (2005). Identifying hallmarks of consciousness in non-mammalian species. Consciousness and cognition, 14(1), 169–187.
· Edelman, D. B., & Seth, A. K. (2009). Animal Consciousness: A Synthetic Approach. Trends in Neuroscience, 32(9), 476–484.
· Elwood, R. W., & Appel, M. (2009). Pain experience in hermit crabs? Animal Behaviour, 77, 1243–1246.
· Epstein, R., & Lanza, R. P., & Skinner, B. F. (1981). Self-awareness in the pigeon. Science, 212, 695–696.
· Farah, M. J. (2008). Neuroethics and the Problem of Other Minds: Implications of Neuroscience for the Moral Status of Brain-Damaged Patients and Nonhuman Animals. Neuroethics, 1, 9–18.
· Feeney, M., & Roberts, W., & Sherry, D. (2011). Black-Capped Chickadees (Poecile artricapillus) Anticipate Future Outcomes of Foraging Choices. Journal of Experimental Psychology, 37(1), 30–40.
· Feinberg, T. E., & Mallatt, J. (2013). The evolutionary and genetic origins of consciousness in the Cambrian Period over 500 million years ago. Frontiers in Psychology, 4, 00667.
· Fisher, J. A. (1990). The Myth of Anthropomorphism. In M. Bekoff & D. Jamieson (eds.), Interpretation and explanation in the study of animal behavior: Interpretation, intentionality, and communication (Vol. 1) (pp. 96–116). Boulder: Westview Press.
· Fitzpatrick, S. (2008). Doing Away with Morgan's Canon. Mind & Language, 23(2), 224–226.
· Froese, T., & Gould, C., & Seth, A. K. (2011). Validating and calibrating first-and second-person methods in the science of consciousness. Journal of Consciousness Studies, 18(2), 38–64.
· Fujita, K., & Blough, D. S., & Blough, P. M. (1991). Pigeons see the Ponzo illusion. Animal Learning and Behavior, 19, 283–293.
· Gaita, R. (2003). The Philosopher's Dog: Friendships with Animals. London: Routledge.
· Gallagher, S. (2008). Direct perception in the intersubjective context. Consciousness and Cognition, 2, 535–543.
· Gallup, G. G. Jr. (1970). Chimpanzees: Self-Recognition. Science, 167(3914), 86-87.
· ––– (1982). Self‐awareness and the emergence of mind in primates. American Journal of Primatology, 2(3), 237–248.
· ––– (1986). Do minds exist in species other than our own? Neuroscience & Biobehavioral Reviews, 4, 631–641.
· Gallup, G. G. Jr., & Anderson, J. R., & Shillito, D. J. (2002). The Mirror Test. In M. Bekoff, C. Allen & G. Burghardt (eds.), The Cognitive Animal (pp. 325–334). Cambridge, MA: MIT Press.
· Gardner, R. A., & Gardner, B. T., & Van Cantfort, T. E. (1989). Teaching sign language to chimpanzees. Albany, NY: SUNY Press.
· Gennaro, R. J. (2004). Higher-order thoughts, animal consciousness, and misrepresentation: A reply to Carruthers and Levine. In R. Gennaro (ed.), Higher-Order Theories of Consciousness: An Anthology (pp. 45–66). Amsterdam: John Benjamins.
· Ginsburg, S., & Jablonka, E. (2007). The transition to experiencing: I. Limited learning and limited experiencing. Biological Theory, 2(3), 218–230.
· Ginsburg, S., & Jablonka, E. (2007). The transition to experiencing: II. The evolution of associative learning based on feelings. Biological Theory, 2(3), 231–243.
· Giurfa, M., et al. (2001). The concepts of ‘sameness’ and ‘difference’ in an insect. Nature, 410, 930–933.
· Grandin, T. (1995). Thinking In Pictures: and Other Reports from My Life with Autism. New York: Doubleday.
· ––– (2004). Animals in Translation: Using the Mysteries of Autism to Decode Animal Behavior. New York: Scribner.
· Griffin, D. R. (1976). The Question of Animal Awareness: Evolutionary Continuity of Mental Experience. New York: Rockefeller University Press.
· ––– (1978). Prospects for a cognitive ethology. Behavioral and Brain Sciences, 4, 527–38.
· ––– (1984). Animal Thinking. Cambridge, MA: Harvard University Press.
· ––– (1992). Animal Minds. Chicago: University of Chicago Press.
· ––– (2002). What is it like? In M. Bekoff, C. Allen & G. Burghardt (eds.), The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition (pp. 471–474). Cambridge, MA: MIT Press.
· Griffin, D. R. (ed.). (1982). Animal Mind – Human Mind.Berlin: Springer.
· Griffin, D. R., & Speck, G. B. (2004). New evidence of animal consciousness. Animal Cognition, 7, 5–18.
· Güzeldere, G. (1995). Is consciousness the perception of what passes in one's own mind? In T. Metzinger (ed.), Conscious Experience (pp. 335–357). Paderborn: Schöningh/Imprint Academic.
· Hare, B., et al. (2000). Chimpanzees know what conspecifics do and do not see. Animal Behavior, 59, 771–785.
· Hare, B., Call, J., & Tomasello, M. (2001). Do chimpanzees know what conspecifics know? Animal Behaviour, 63, 139–151.
· Hare, B., & Wrangham, R. (2002). Integrating two evolutionary models for the study of social cognition. In M. Bekoff, C. Allen & G. Burghardt (eds.), The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition (pp. 363–370). Cambridge, MA: MIT Press.
· Hauser, M. D., Chomsky, N., & Fitch, W. T. (2002). The Faculty of Language: What is It, Who Has It, and How Did it Evolve? Science, 298(5598), 1569–1579.
· Hauser, M. D., et al. (1995). Self-recognition in primates: phylogeny and the salience of species-typical features. Proceedings of the National Academy of Sciences, 92, 10811–10814.
· Hauser, M. D., et al. (2001). Cotton-top tamarins (Saguinus oedipus) fail to show mirror-guided self-exploration. American Journal of Primatology, 53, 131–137.
· Heyes, C. (1998). Theory of mind in nonhuman primates. Behavioral and Brain Sciences, 21, 101–148.
· Hurley, S., & Nudds, M. (eds.). (2006). Rational Animals? Oxford: Oxford University Press.
· Huxley, T. H. (1874). On the hypothesis that animals are automata, and its history. Fortnightly Review, 22, 199–245.
· Institute for Laboratory Animal Research (2009). Recognition and Alleviation of Pain in Laboratory Animals, Committee Report. Washington, DC: National Research Council.
· Jackson, R., & Wilcox, S. (1993). Observations in nature of detouring behavior by Portia Fimbriata, a web-invading aggressive mimic jumping spider from Queensland. Journal of Zoology, 230, 135–139.
· James, W. (1890). The Principles of Psychology. New York: Dover.
· Jamieson, D. (1998). Science, knowledge, and animal minds. Proceedings of the Aristotelian Society, 98, 79–102.
· Jamieson, D., & Bekoff, M. (1992). Carruthers on nonconscious experience. Analysis, 52, 23–28.
· Kennedy, J. S. (1992). The new anthropomorphism. New York: Cambridge University Press.
· Langford, D., et al. (2006). Social Modulation of Pain as Evidence for Empathy in Mice. Science, 312, 1967–1970.
· Leopold, D. A., & Logothetis, N. K. (1996). Activity changes in early visual cortex reflect monkeys' percepts during binocular rivalry. Nature, 379, 549–553.
· Leopold, D. A., Maier, A., & Logothetis, N. K. (2003). Measuring Subjective Visual Perception in the Nonhuman Primate. Journal of Consciousness Studies, 10, 115–130.
· Lewis, C. S. (2009). The problem of pain. London: HarperCollins e-book.
· Lorenz, K. (1971). Comparative studies of the motor patterns of Anatinae. In K. Lorenz (ed.), Studies in Animal and Human Behavior (Vol. 2) (pp. 14–114). Cambridge, Mass: Harvard University Press.
· ––– (1971). Comparative studies of the motor patterns of Anatinae. In K. Lorenz (ed.), Studies in Animal and Human Behavior (Vol. 2) (pp. 14–114). Cambridge, MA: Harvard University Press.
· ––– (1971). Do animals undergo subjective experiences? In K. Lorenz (ed.), Studies in Animal and Human Behavior (pp. 323–337). Cambridge, MA: Harvard University Press.
· Lurz, R. W. (2011). Mindreading Animals: The Debate Over What Animals Know About Other Minds. Cambridge, MA: MIT Press.
· Lycan, W. G. (1995). Consciousness as Internal Monitoring, I: The Third Philosophical Perspectives Lecture. Philosophical Perspectives, 9, 1–14.
· ––– (1996). Consciousness and Experience. Cambridge, MA: MIT Press.
· Margulis, L. (2001). The conscious cell. Annals of the New York Academy of Sciences, 929, 55–70.
· Mather, J. A. (2008). Cephalopod consciousness: behavioral evidence. Consciousness and Cognition, 17, 37–48.
· Mathews, K. (2008). Neuropathic Pain in Dogs and Cats: If Only They Could Tell Us If They Hurt. In K. Mathews (ed.), Update on Management of Pain, An Issue of Veterinary Clinics: Small Animal Practice (pp. 1365–1414). Philadelphia, PA: Saunders.
· Melzack, R., & Wall, P. (1965). Pain mechanisms: a new theory. Science, 150, 971–9.
· Mendl, M., Paul, E. S., & Chittka, L. (2011). Animal behaviour: emotion in invertebrates? Current Biology, 12, R463-R465.
· Merker, B. (2005). The liabilities of mobility: A selection pressure for the transition to consciousness in animal evolution. Conscious and Cognition, 14(1), 89–114.
· Mitchell, R. W. (2002). Kinesthetic visual matching, imitation, and self-recognition. In M. Bekoff, C. Allen & G. Burghardt (eds.), The Cognitive Animal (pp. 345–351). Cambridge, MA: MIT Press.
· Mogil, J. S. (2009). Animal models of pain: progress and challenges. Nature Reviews: Neuroscience, 10, 283–294.
· Morgan, C. L. (1894). An Introduction to Comparative Psychology. New York: Scribner.
· Myserson, J., Miezin, F. M., & Allman, J. M. (1981). Binocular rivalry in macaque monkeys and humans: a comparative study in perception. Behaviour Analysis Letters, 1, 149–159.
· Nagel, A. H. M. (1997). Are Plants Conscious? Journal of Consciousness Studies, 4(3), 215–230.
· Nishihara, H., Hasegawa, M., & Okada, N. (2006). Pegasoferae, an unexpected mammalian clade revealed by tracking ancient retroposon insertions. Proceedings of the National Academy of Sciences, 26, 9929–9934.
· Olson, R. (1990). Science Defied and Science Deified: The Historical Significance of Science in Western Culture. Berkeley and Los Angeles, California: University of California Press.
· Panksepp, J. (2004). Affective Neuroscience: The Foundations of Human and Animal Emotions. New York: Oxford University Press.
· ––– (2005). Affective consciousness: Core emotional feelings in animals and humans. Consciousness and Cognition, 14, 30–80.
· Panksepp, J., & Burgdorf, J. (2003). ‘Laughing’ rats and the evolutionary antecedents of human joy? Physiology and Behavior, 79, 533–47.
· Pepperberg, I. M. (1999). The Alex Studies: Cognitive and Communicative Abilities of Grey Parrots. Cambridge, MA: Harvard University Press.
· Plotnik, J. M., Waal, F., & Reiss, D. (2006). Self-Recognition in an Asian Elephant. Proceedings of the National Academy of Sciences, 103, 17053–17057.
· Povinelli, D. J. (1996). Chimpanzee theory of mind?: the long road to strong inference. In P. Carruthers & P. Smith (eds.), Theories of Theories of Mind (pp. 293–329). Cambridge: Cambridge University Press.
· Povinelli, D. J., & Giambrone, S. J. (2000). Inferring Other Minds: Failure of the Argument by Analogy. Philosophical Topics, 27, 161–201.
· Preston, S. D., & de Waal, F. B. M. (2002). Empathy: Its ultimate and proximate bases. Behavioral and Brain Sciences, 25, 1–72.
· Price, D. (2000). Psychological and neural mechanisms of the affective dimension of pain. Science, 288, 1769–72.
· Prinz, J. (2005). A Neurofunctional Theory of Consciousness. In A. Brook & K. Akins (eds.), Cognition and the Brain: The Philosophy and Neuroscience Movement (pp. 381–96). New York: Cambridge University Press.
· Prior, H., Schwartz, A., & Güntürkün, O. (2008). Mirror-Induced Behavior in the Magpie (Pica pica): Evidence of Self-Recognition. Public Library of Science/Biology, 6(8), e202.
· Radick, G. (2000). Morgan's canon, Garner's phonograph, and the evolutionary origins of language and reason. British Journal for the History of Science, 33, 3–23.
· Radner, D. (1994). Heterophenomenology: learning about the birds and the bees. Journal of Philosophy, 91, 389–403.
· Radner, D., & Radner, M. (1986). Animal Consciousness. Amherst, NY: Prometheus Books.
· Rajala, A. Z., et al. (2010). Rhesus Monkeys (Macaca mulatta) Do Recognize Themselves in the Mirror: Implications for the Evolution of Self-Recognition. PLoS ONE, 5(9), e12865.
· Rees, G., Kreiman, G., & Koch, C. (2002). Neural correlates of consciousness in humans. Nature Reviews Neuroscience, 3, 261–270.
· Regan, T. (1983). The Case for Animal Rights. Berkeley: University of California Press.
· Reiss, D., & Marino, L. (2001). Mirror Self-Recognition in the Bottlenose Dolphin: A Case of Cognitive Convergence. Proceedings of the National Academy of Sciences, 98(10), 5937–5942.
· Roberts, W. (2012). Evidence for future cognition in animals. Learning and Motivation, 43, 169–180.
· Robinson, W. (2007). Evolution and epiphenomenalism. Journal of Consciousness Studies, 11, 27–42.
· Rochat, P., & Zahavi, D. (2011). The uncanny mirror: A re-framing of mirror self-experience. Consciousness and cognition, 2, 204–213.
· Rollin, B. E. (1989). The Unheeded Cry: Animal Consciousness, Animal Pain and Science. New York: Oxford University Press.
· Romanes, G. (1882). Animal Intelligence. London: Routledge & Kegan Paul.
· Rose, J. D. (2002). The neurobehavioral nature of fishes and the question of awareness and pain. Reviews in Fisheries Science, 10, 1–38.
· Rosenthal, D. (1986). Two concepts of consciousness. Philosophical Studies, 49, 329–359.
· ––– (1993). Thinking that one thinks. In M. Davies & G. Humphreys (eds.), Consciousness (pp. 197–223). Oxford: Blackwell.
· ––– (1993). Thinking that one thinks. In M. Davies & G. Humphreys (eds.), Consciousness (pp. 197–223). Oxford: Blackwell.
· Savage-Rumbaugh, S., & Lewin, R. (1996). Kanzi: The Ape at the Brink of the Human Mind. New York: John Wiley & Sons.
· Searle, J. (1998). Animal minds. Etica & Animali, 9, 37–50.
· Seth, A., Baars, B. J., & Edelman, D. B. (2005). Criteria for consciousness in humans and other mammals. Consciousness and Cognition, 14, 119–139.
· Shriver, A. (2006). Minding Mammals. Philosophical Psychology, 19, 433–442.
· ––– (2009). Knocking Out Pain in Livestock: Can Technology Succeed Where Morality has Stalled? Neuroethics, 2(3), 115–124.
· Shumaker, R. W., & Swartz, B. (2002). When traditional methodologies fail: Cognitive studies of great apes. In M. Bekoff, C. Allen & G. Burghardt (eds.), The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition (pp. 335–43). Cambridge, MA: MIT Press.
· Singer, P. (1990 [1975]). Animal Liberation. New York: Avon Books.
· Skinner, B. F. (1953). Science and Human Behavior. New York: Macmillan.
· Smith, J., & Boyd, K. (eds.). (1991). Lives in the Balance: The Ethics of Using Animals in Biomedical Research.New York: Oxford University Press.
· Smith, J. D. (2009). The study of animal metacognition. Trends in Cognitive Sciences, 13(9), 389–396.
· Smith J. D., Shields, W. E., & Washburn D. S. (2003). The comparative psychology of uncertainty monitoring and metacognition. Behavioral and Brain Sciences, 26, 317–373.
· Sneddon, L. U., Braithwaite, V. A., & Gentle, M. J. (2003). Do fish have nociceptors: evidence for the evolution of a vertebrate sensory system. Proceedings of the Royal Society London B, 270, 1115–1121.
· Sober, E. (1998). Morgan's Canon. In C. Allen & D. Cummins (eds.), The evolution of mind (pp. 224–242). Oxford: Oxford University Press.
· ––– (2000). Evolution and the problem of other minds. Journal of Philosophy, 97, 365–386.
· ––– (2005). Comparative psychology meets evolutionary biology: Morgan's canon and cladistic parsimony. In L. Datson & G. Mitman (eds.), Thinking with Animals: New Perspectives on Anthropomorphism (pp. 85–99). New York: Columbia University Press.
· ––– (2012). Anthropomorphism, Parsimony, and Common Ancestry. Mind & Language, 27(3), 229–238.
· Soley, F. G., & Alvarado-Díaz, I. (2011). Prospective thinking in a mustelid? Eira barbara (Carnivora) cache unripe fruits to consume them once ripened. Naturwissenschaften, 8, 693–698.
· Sorabji, R. (1993). Animal Minds and Human Morals: the origins of the Western debate. Ithaca, NY: Cornell University Press.
· Spalding, D. A. (1872). Instinct: With original observations on young animals. MacMillan's Magazine, 27, 283–93.
· Srinivasan, M. V. (2010). Honey Bees as a Model for Vision, Perception, and Cognition. Annual Review of Entomology, 55, 267–284.
· Steiner, G. (2008). Animals and the Moral Community: Mental Life, Moral Status, and Kinship. New York: Columbia University Press.
· Stoerig, P., & Cowey, A. (1997). Blindsight in man and monkey. Brain, 120, 535–559.
· Stoerig, P., Zontanou, A., & Cowey, A. (2002). Aware or Unaware: Assessment of Cortical Blindness in Four Men and a Monkey. Cerebral Cortex, 12(6), 565–574.
· Suddendorf, T., & Corballis, M. C. (1997). Mental time travel and the evolution of the human mind. Genetic, social, and general psychology monographs, 123(2), 133–167.
· ––– (2007). The evolution of foresight: What is mental time travel, and is it unique to humans? Behavioral and Brain Sciences, 3, 299–312.
· Sufka, K. J., Weldon, M., & Allen, C. (2009). The Case for Animal Emotions: Modeling Neuropsychiatric Disorders. In J. Bickle (ed.), The Oxford Handbook of Philosophy of Neuroscience (pp. 522–536). New York: Oxford University Press.
· Terrace, H. S., & Son, L. K. (2009). Comparative metacognition. Current Opinion in Neurobiology, 19(1), 67–74.
· Thomas, R. K.(2001). Lloyd Morgan's canon: A history of misrepresentation. Retrieved from: http://htpprints.yorku.ca/archive/00000017 on June 8, 2006.
· Thorndike, E. L. (1911). Animal Intelligence. Darien, CT: Hafner.
· Tononi, G. (2008). Consciousness as integrated information: a provisional manifesto. The Biological Bulletin, 215(3), 216–242.
· Trestman, M. (2013). The Cambrian explosion and the origins of embodied cognition. Biological Theory, 8(1), 80–92.
· ––– (2013). The modal breadth of consciousness. Philosophical Psychology, 27(6), 843-861.
· Trout, J. D. (2001). The biological basis of speech: What to infer from talking to the animals. Psychological Review, 108(3), 523–549.
· Tulving, E. (1985). Memory and consciousness. Canadian Psychology/Psychologie Canadienne, 26(1), 1–12.
· Tye, M. (2000). Consciousness, Color, and Content. Cambridge, MA: MIT Press.
· Varner, G. (1998). In Nature's Interests? New York: Oxford University Press.
· ––– (2012). Personhood, Ethics, and Animal Cognition: Situating Animals in Hare's Two Level Utilitarianism. New York: Oxford University Press.
· Velmans, M. (2012). The evolution of consciousness. Contemporary Social Science, 7(2), 117–138.
· Wallace, A. R. (1867). The philosophy of birds' nests. Intellectual Observer, 11, 413–20.
· Walters, E. T. (1996). Comparative and Evolutionary Aspects of Nociceptor Function. In C. Belmonte & F. Cervero (eds.), Neurobiology of Nociceptors (pp. 92–114). New York: Oxford University Press.
· Watson, J. B. (1928). The Ways of Behaviorism. New York: Harper.
· White, G. (1789). The Natural History of Selbourne. London and New York: Dent/Dutton.
· Wilcox, R. S., & Jackson, R. R. (1998). Cognitive abilities of araneophagic jumping spiders. In C. Kamil (ed.), Animal cognition in nature: the convergence of psychology and biology in laboratory and field. Academic (pp. 411–434). San Diego: Academic Press.
· Wilkes, K. (1984). Is consciousness important? British Journal for the Philosophy of Science, 35, 223–243.
· Wilson, M. D. (1995). Animal ideas. Proceedings and Addresses of the American Philosophical Association, 69, 7–25.
· Wynne, C. (2004). Do Animals Think? Princeton, NJ: Princeton University Press.
· Zahavi, D. (2011). Empathy and direct social perception. Review of Philosophy and Psychology, 2(3), 541–558.
.jpg)








