
.jpg)




Биология развития
Впервые опубликовано 23 марта 2015 г.
Биология развития — это наука, объясняющая то, как множество различных взаимодействующих процессов порождает гетерогенные формы, определяет размер и структурные особенности организма, возникающие на стадиях от эмбриона до взрослой особи, или, в более общем смысле, на протяжении всего жизненного цикла. Она представляет образцовую область современной экспериментальной биологии, фокусирующуюся на феноменах, занимавших натурфилософов и ученых на протяжении более двух тысячелетий. Философы биологии проявили высокий интерес к биологии развития из-за потенциальной значимости развития для понимания эволюции, темы редукционизма в генетических объяснениях, а также возросшего внимания к отдельным элементам конкретных исследовательских программах. Биология развития демонстрирует богатый массив материальных и концептуальных практик, анализ которых может позволить лучше понять представленные в экспериментальной науке о жизни способы научного теоретизирования. В данной статье кратко рассматриваются некоторые ключевые явления онтогенеза, а затем исследуются четыре области, важные и перспективные для рефлексии над эпистемологией биологии развития.
Обзор
Исторические соображения
Биология развития – это наука, объясняющая, как множество различных взаимодействующих процессов порождает гетерогенные формы, определяет размеры и структурные особенности организма, возникающие на стадиях от эмбриона до взрослой особи, или, в более общем смысле, на протяжении всего жизненного цикла (Love 2008b; Minelli 2011a). Она представляет одну из важнейших областей современной экспериментальной биологии, фокусирующуюся на феноменах, занимавших натурфилософов и ученых на протяжении более двух тысячелетий. Как динамические отношения между однородными с виду компонентами на ранних стадиях существования эмбриона формируют единый цельный организм, содержащий гетерогенные части в соответствующем расположении и с правильными взаимосвязями? Коротко говоря, как мы можем объяснить онтогенез (или, выражаясь более архаично, возникновение)? В своей книге «О возникновении животных» Аристотель предложил первое систематическое исследование феноменов развития и наметил ключевые проблемы, связанные с появлением иерархически организованных частей и их взаимоотношений (например, между костью и анатомическими особенностями, включающими в себя кость); также он указал на трудность в определении того, как морфологической форме удается надежно закрепиться в потомстве (например, как закрепляется типическая форма и структура придатков у конкретного вида). Возникновение оставалось острым вопросом в течение Раннего Нового Времени и изучалось многими крупнейшими мыслителями, в числе которых были Уильям Гарвей, Рене Декарт, Роберт Бойль, Пьер Гассенди, Николас Мальбранш, Готфрид Вильгельм Лейбниц, Энн Конвей, Иммануил Кант и другие (Smith 2006). Наблюдения за изменениями в ходе жизненного цикла (например, метаморфозами) поддерживали эти усилия и приводили к сенсационным выводам, вроде отрицания (в случае Лейбница) самого возникновения sensu stricto [в узком смысле].
Основной темой, оформившейся в этой истории исследования, стало различие между эпигенезом и преформизмом (см. статью epigenesis and preformationism). Сторонники теории эпигенеза утверждали, что гетерогенные, сложные особенности формы возникают из однородных, менее сложных эмбриональных структур через воздействующие друг на друга процессы. Соответственно, объяснение онтогенеза этих особенностей формы требует учитывать протекание этих взаимодействий. Сторонники теории преформации, в свою очередь, утверждали, что сложная форма предсуществует у эмбрионов и «разворачивается» через обычные процессы роста. Полное объяснение, в таком случае, включает в себя подробный анализ протекания роста. Хотя преформизм берет на себя более легкую (в плане объяснения) задачу описания возникновения формы в процессе онтогенеза (если мы исходим из предположения, что рост объяснить легче, чем взаимодействия процессов), он также должен объяснить, как формируется отправная точка следующего поколения, обладающая требуемыми разнородными сложными особенностями. Иногда исследователи, отвечая на этот вопрос, изображали этот процесс формирования так, будто организм заключает внутри себя бесконечное количество своих все более или более маленьких миниатюр (рис. 1). Ответы, которые предлагала теория эпигенеза, часто опирались на формы телеологического мышления (см. статью teleological notions in biology) в попытках объяснить, почему взаимодействия между однородными компонентами в конечном итоге приводят к сложному интегрированному целому – организму. Хотя ничто не мешает совмещать черты этих двух подходов к объяснению различных аспектов развития, они часто противопоставлялись друг другу, выступая в качестве дихотомической пары (Rose 1981; Smith 2006).
Рисунок 1: сделанное в период Раннего Нового Времени (в качестве иллюстрации преформизма) изображение крошечного человечка внутри семени.
В конце XIX и начале XX веков тема развития занимала особое место в дебатах вокруг витализма, как, например, разногласие между Вильгельмом Ру и Гансом Дришем по поводу правитльного объяснения онтогенеза (Maienschein 1991). Ру считал, что оплодотворенная яйцеклетка содержит унаследованные элементы, представляющие собой разные характеристики организма. Во время процесса клеточного деления эти элементы распределяются между дочерними клетками неравномерно, наделяя их разной судьбой. Дриш, напротив, полагал, что каждая клетка в процессе деления сохраняет весь свой потенциал даже несмотря на произошедшую дифференциацию. Хотя данный сюжет часто описывается через метафизику жизни (витализм vs. материализм), предложенная Дришем интерпретация развития и автономии организма имела также эпистемологические аспекты (Maienschein 2000). Разногласия относительно способов объяснения порождали разнородные экспериментальные подходы и противоположные взгляды на саму природу дифференциации на ранней стадии онтогенеза (например, на то, в какой степени клетки предварительно заданы). Хорошо известная в философии тема, фигурирующая в этих дискуссиях, как эпистемологических, так и метафизических, – это статус редукционизма в биологии (см. reductionism in biology). С середины XX века эмбриология – научная дисциплина, изучающая развитие – постепенно превращалась в биологию развития со множеством как переработанных, так и неизменных элементов (Berrill 1961). Наряду с проблемой редукционизма ключевым аспектом данного процесса выступает экспансия молекулярных методов в экспериментальной (в отличие от сравнительной) эмбриологии (Fraser and Harland 2000) – одновременно с указанием на то, что гены служат важным источником объяснения (см. Раздел 3.1 ). Эта сложная и увлекательная история была подробно описана в других работах (см., например, Oppenheimer 1967, Horder et al 1986, Hamburger 1988, Maienschein et al., 2005, Gilbert 1991; проект Embryo).
Биология развития не была основной сферой исследований в философии биологии (см. philosophy of biology), хотя за последние два десятилетия эта ситуация изменилась – особенно в силу потенциальной значимости развития для понимания эволюции (Love 2015; раздел 5), темы редукционизма в биологии и объяснений средствами молекулярной генетики (Robert 2004, Rosenberg 2006, Section 3), а также возросшего внимания к деталям конкретных исследовательских программ, например, биологии стволовых клеток (Fagan 2013). Не следует, впрочем, забывать, что в XX онтогенез веке был предметом интереса ряда философов, как видно из предложенного Эрнестом Нагелем анализа иерархической организации и редукции в развитии живых систем (Nagel 1961: 432ff). Биология развития предоставляет современной философии науки богатый набор материальных и концептуальных практик, анализ которых позволяет лучше понять применяющиеся в экспериментальной науке о жизни типы научного теоретизирования. В данной статье кратко рассматриваются некоторые ключевые явления онтогенеза, а затем исследуются четыре области, важные и перспективные для рефлексии над эпистемологией биологии развития.
Явления развития
Биология развития – это наука, стремящаяся объяснить, как структура организмов изменяется со временем. Структура, которую также можно назвать морфологией или анатомией организма, охватывает расположение его частей, количество частей и их различные типы. (Slack 2006: 6)
В учебниках (например, Gilbert 2010, Slack 2013, Wolpert et al. 2010) обычно приводится хрестоматийный набор происходящих с изменяющимися структурами событий, которые развертываются в процессе развития животного. Первым из них является оплодотворение (у видов, воспроизводящихся половым путем), где уже полуорганизованная яйцеклетка сливается с клеткой спермы, после чего происходит слияние их ядер – оно позволяет достичь взаимодополнения их общего генетического материала. На втором шаге оплодотворенная яйцеклетка проходит через несколько циклов дробления, которые представляют собой митотические деления без роста клеток, в ходе которых зигота делится на много отдельных клеток (рис. 2). После множества циклов расщепления получившийся сферический конгломерат клеток (называемый бластулой) начинает демонстрировать определенную спецификацию зародышевых слоев (энтодермы, мезодермы и эктодермы), а затем переходит к выпячиванию с одного конца: это начало комплексного процесса, называемого гаструляцией, который в конечном итоге формирует сквозную кишку. Все три зародышевых слоя, из которых потом возникнут клетки конкретных типов (например, как нервные клетки из эктодермы) окончательно складываются во время гаструляции или вскоре после её завершения. Органогенезом называется формирование тканей и органов посредством взаимодействия и перестановок клеточных групп. События, ограниченные отдельными таксономическими группами, включают нейруляцию у хордовых, тогда как другие события соответствуют способу развития (метаморфоза личинки до взрослой особи) или связаны с индивидуальной травмой (регенерация конечности).
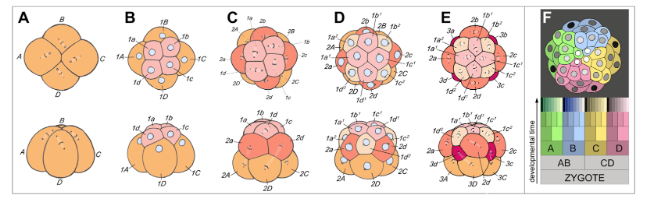
Рисунок 2: Пример эмбрионального дробления у эмбрионов морских улиток, показывающий судьбу различных клеточных линий в течение времени их развития.
В основе этих различных событий развития и возникающих в итоге особенностей формы (например, сквозной кишки, образованной после гаструляции, или сердца, образовавшегося во время органогенеза) лежат несколько ключевых процессов. Они имеют решающее значение для онтогенеза формы (form) и напрямую связаны с основными исследовательскими вопросами в биологии развития (раздел 2). Во-первых, клеточные свойства – такие, как ее форма (shape), – изменяются во время онтогенеза: посредством функции дифференциации клетки получают конкретные судьбы, включая изменение их вида (рисунок 3). Во-вторых, области клеток в эмбрионе обозначаются на основе изменений их расположения и построения: эти изменения соответствуют различным осям в разных частях эмбриона (например, дорзально-вентральной, передне-задней, лево-правой и проксимально-дистальной). Последовательное складывание этих областей называется формированием паттернов. В-третьих, клетки проходят транслокацию и собираются в слои (например, эндодерму и эктодерму, за которыми во многих клеточных родословных следует мезодерма), а затем и ткани (агрегации дифференцированных типов клеток). В-четвертых, клетки и ткани мигрируют и взаимодействуют, производя новые компоновки и формы (shape), состоящие из нескольких слоев тканей с новыми функциями (т.е. органами). Последние два набора процессов обычно называют морфогенезом (Davies 2005); они происходят посредством множества различных механизмов. В-пятых, у особи увеличивается размер различных особенностей формы, что заметно при сравнении зиготы со взрослым организмом, хотя не менее поразительно и изменение пропорций между различными особенностями организма (аллометрия).
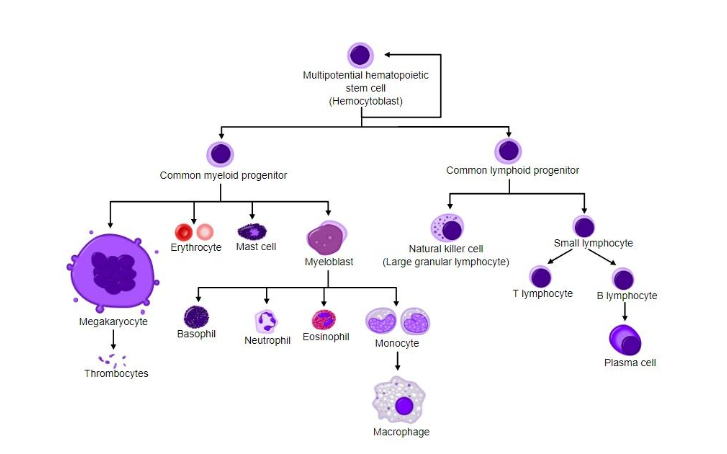
Рисунок 3: Простая иллюстрация типов дифференциации, связанных с обнаруженными в крови клеточными компонентами.
Ни один из этих процессов не протекает изолированно, а объяснения конкретных особенностей формы обычно основываются одновременно на нескольких из них. Тем самым предполагается, что другие признаки возникли в онтогенезе ранее, в ходе спецификации заданных характеристик и комбинаций различных процессов. Все это ставит широкий круг исследовательских задач: как различные итерации и комбинации упомянутых процессов порождают особенности формы в ходе онтогенеза? Более отчетливо это можно увидеть на примере кардиогенеза позвоночных. Как сердце позвоночных с его внутренними и внешними формой и структурой возникает в ходе онтогенеза (Harvey 2002)? Как сердце начинает проявлять свою лево/право-стороннюю асимметрию в грудной полости тела? Что заставляет клетки принимать судьбу мышечных клеток или других определенных тканей, чтобы взаимодействовать в предполагаемом регионе сердца? Как мышечные клетки мигрируют, объединяются и дифференцируются в правильном месте? Как внутренняя часть сердца принимает специфическую трубчатую структуру с различными камерами (которая отличается у разных видов позвоночных)? Как сердце растет с определенной скоростью и достигает определенного размера? Решения, подходящие для объяснения онтогенеза формы, характеризуют каузальные факторы, ответственные за то, как протекают различные процессы и как в итоге они приводят к различным результатам (Раздел 3 ).
Эпистемологическая организация биологии развития
На протяжении долгой истории исследований процессов развития снова и снова заявляла о себе идея о том, что объяснение онтогенеза формы состоит из множества взаимосвязанных вопросов о различных явлениях (см. раздел 1.2). Иногда философы пытались свести все эти вопросы к одной широкой проблеме.
Подлинный вопрос, касающийся онтогенеза многоклеточных, состоит в следующем: как одна клетка порождает необходимое количество дифференцированных клеточных родословных со всеми их правильными, ведущими к развитию взаимодействиями, необходимыми для воспроизведения формы зрелого организма (Moss 2002: 97).
Главная задача, стоящая перед биологией развития, заключается в том, чтобы понять, как относительно простая и однородная клеточная масса может дифференцироваться в относительно сложный и гетерогенный организм, близкий к его предшественнику(-ам) в соответствующих аспектах (Robert, 2004: 1).
Подобная формулировка не является с необходимостью неверной, но ее использование может привести к искажениям. Например, Филип Китчер утверждал, что:
В современной биологии развития существует ... неуверенность по поводу того, как ответить на большой и размытой вопрос: "как развиваются организмы (Kitcher, 1993: 115)
Данное утверждение попросту неверно. Хотя верно, что эти вопросы появлялись с различной частотой и настойчивостью в течение всей истории дисциплины, а способность отвечать на них (равно как и характер самих вопросов) зависела от различных стратегий и методов исследований, приведенный выше вопрос не был самой большой и самой главной проблемой. Впрочем, тщательное изучение структуры вопросов биологии развития – не просто упражнение в разъяснении. Крайне важно понять, как организована наука биологии развития.
Теории развития нет?
Хотя в философии научные дисциплины часто ассоциируют с теориями – как если бы индивидуализация науки зависела от некой определяющей теории или группы моделей – биологию развития редко представляют через некую теорию развития. Вместо этого мы обнаруживаем ссылки на семейства подходов (генетика развития, экспериментальная эмбриология, клеточная биология и молекулярная биология) или каталоги «ключевых молекулярных компонентов» (семейства факторов транскрипции, индуцирующие факторные группы, цитоскелет или молекулы адгезии клеток и компоненты внеклеточного матрикса). В основных учебниках (например, Slack 2013, Wolpert и др., 2010; Gilbert 2010) нет никакой стандартной теории или группы моделей, которые бы предоставляли некий теоретический каркас. Отсутствие каких-либо ссылок на теорию развития или хотя бы на некоторый набор основных объяснительных моделей prima facie [на первый взгляд] озадачивает. Почему же так сложно определить определяющую теорию биологии развития? Здесь есть три возможных интерпретации. Во-первых, отсутствие ссылок на теорию не указывает на отсутствие теории; можно реконструировать теорию (или теории) биологии развития из соответствующего дискурса (например, указывая на множество схожих молекулярных моделей). Вторая интерпретация заключается в том, что отсутствие ссылки на теорию указывает на незрелость биологии развития. Поскольку зрелые науки всегда имеют систематические теории, мы должны стремиться сформулировать таковую и для области исследований онтогенеза. Третья интерпретация заключается в том, что отсутствие ссылки на общую теорию следует принимать как данность.
Биология развития не является незрелой наукой, которая пытается хоть как-то объяснить изучаемые ей явления: «некоторые из основных процессов и механизмов эмбрионального развития в настоящее время поняты достаточно хорошо» (Slack 2013: 7). Скорее, к этому варианту интерпретации подталкивает концепция об обязательствах зрелой науки, предполагающая, что теории – это абстрактные системы с небольшим набором законов или основных принципов. С другой стороны, мы можем предположить, что биология развития уже имеет теорию в другом обличье, такую, на которую биологи развития не ссылаются как на теорию. Эта версия возникает из родственной концепции науки (научные дисциплины должны иметь теории), к которой прибавляется допущение о том, что структура теории может пониматься различным образом, например, как сочетания моделей без основополагающих законов, несмотря на предположение, что теория играет аналогичную организационную роль в управлении ходом исследования. Однако такое предположение следует оспорить и отклонить в случае биологии развития на методологических основаниях. Анализ теоретизирования в науке должен работать в условиях эпистемической прозрачности и не постулировать «скрытую» структуру рассуждений (Love 2012). Этот критерий основан на предпосылке, что исследователям, включенным в научную практику (т.е. ученым), должен быть доступен некий минимум успеха в научном исследовании. Если мы постулируем не присутствующую в научном дискурсе скрытую структуру для обоснования индуктивных умозаключений, объяснений или других форм теоретизирования, то мы рискуем упустить из виду то, как сами ученые получают доступ к этой структуре для ее оценки (Woodward 2003: ch. 4). В результате, если смотреть с точки зрения участников, успехи наук становятся таинственными.
Эпистемическая прозрачность требует дескриптивного соответствия между философскими представлениями о науке и научной практикой. Это не означает, что любые претензии любого ученого должны приниматься с одинаковым доверием. Скорее, главное здесь – это проявляющиеся повсюду особенности практики. Проблема допущения, что при объяснении необходимо указывать на некие законы, состоит в том, такие законы просто-напросто отсутствуют во многих успешных науках, регулярно производящих объяснения, нежели в том, что ни один ученый никогда не обращается к законам как к источнику объяснения. Проявляющиеся повсюду особенности научных практик должны быть четко обозначены в философских описаниях науки. Вот почему не кажется удивительным, что стремление к построению теории присутствует и у некоторых биологов развития: «Разработка теории имеет первостепенное значение для любой дисциплины» (Sommer, 2009: 417). Однако тот факт, что подобные призывы редки, означает, что мы не должны считать теории действительно необходимыми для направления и организации исследований в конкретной области знания. [ 3 ]
Когда-то считалось, что каждая наука, чтобы иметь объяснительную силу, должна располагать некими законами, однако теперь это не считается необходимым (Giere 1999; Woodward 2003). Ожидание, что в науке теория организовывает и направляет исследования, в сущности, сходно с предыдущим. Прежде всего, оно проистекает из интуитивного предположения о том, чем вообще является зрелая наука. Даже если мы найдем эмпирически успешные и последовательные традиции исследований, которые, однако, не покоятся некой на систематической теоретической основе, эта наука, согласно данной точке зрения, не может считаться зрелой. Можно было бы отказаться от этих квазипозитивистских апелляций к зрелости, прибегая к более гибким интерпретациям теории и ее структуры. Но почему мы вообще полагаем, что теории должны выполнять одни и те же эпистемические задачи? Данное представление о структуре знания является предвзятым и оказывается неправдоподобным в свете разнообразия исследовательских практик в существующих научных областях. Те немногие ученые, которые придерживаются данной философской позиции, руководствуются различными мотивами. Вместо «зрелости» они полагаются на другие основания, например, стремление найти ориентир в ситуации наличия множества беспорядочных биохимических деталей, а также необходимость объединить теорию эволюции с теорией развития.[ 4 ]
Биологи развития признают, что «проклятие деталей» – это та цена (среди многих других), которую приходится платить за стремительный успех биологии развития в течение последних трех десятилетий: «Главный вызов сегодня – экспоненциально возрастающее число деталей» (Slack 2013: ix). Хотя что-то и должно обеспечивать организацию и направление биологии развития, эту задачу не обязательно должны брать на себя теории. Что касается призывов к синтезу эволюции и развития, то за ними часто стоит допущение, что наличие теории развития само является предварительным условием объединения (Sommer 2009): «Наши проблемы ... проистекают из того, что у нас все еще отсутствует отчетливая теория развития» (Minelli 2011a: 4). Однако эта линия аргументации опирается на то, что эволюционная теория обладает выраженной структурой, к чему также должна стремится биология развития. Реальная же научная практика, связанная с эволюционной теорией, демонстрирует более гибкое строение, которое, как хамелеон, быстро адаптируется к разнообразным целям и проектам исследователей эволюции (Love 2010b, 2013). Таким образом, далеко не очевидно, что эволюционную теорию можно считать подходящим образцом. Чтобы продвинуться вперед, было бы продуктивно отказаться от предварительных ожиданий, будто науки должны иметь некие особые теории, управляющие и организующие их деятельность. Вместо этого следует изучать те науки, которые успешно достигают своих целей и приносят значимые результаты, чтобы выяснить, какие именно их черты способствуют их успеху, не предполагая при этом, что они будут одинаковыми для всех наук: «Науку не следует рассматривать в таких терминах и, по сути, ее следовало бы рассматривать иначе» (Giere 1999: 4).
Проблематическая организация
Критерий эпистемической прозрачности (раздел 2.1) заставляет нас подробнее исследовать третий способ интерпретации, опирающийся на тезис, что отсутствие ссылки на теории следует принимать как данность. В первую очередь биология развития организовывается через стабильные, широкие проблемные области, которые соответствуют абстрактным представлениям об основных онтогенетических процессах (дифференциации, формировании паттернов, росте и морфогенезе; раздел 1.2). Но как тогда интерпретировать «теоретические» аспекты биологии развития (например, модели позиционной информации в формирования паттернов) и использование теорий из других областей (например, биохимии)?
Один из способов устранить это затруднение – провести различие между теоретико-информированной наукой (использующей теоретические знания) и теоретико-ориентированной наукой (имеющей теорию, которая направляет исследования и организует знания (Waters 2007b)); биология развития тогда является теоретико-информированной, но не теоретико-ориентированной.
Теории не обязательно должны быть полностью исключены из биологии развития, но, если они и присутствуют, их роль сильно отличается от той, которую приписывают ей стандартные философские объяснения. Биология развития использует теоретические знания в области биохимии, когда обращается к градиентам морфогена для объяснения сегментов тела, или к химической термодинамике, когда обращается к механизмам реакции-диффузии для объяснения паттернов пигментации. Последняя также использует такие полученные из биологии развития теоретические знания как модели позиционной информации.
Биология развития привлекает различные типы теорий, но они не организуют исследования – они не требуются для структурирования знаний и непосредственных исследовательских практик. Биологи развития не сосредоточены на подтверждении и расширении теории механизмов реакции-диффузии, равно они не организуют свое исследование вокруг позиционной информации.[ 5 ] Это теоретическое знание используется при построении объяснений, но оно не дает рекомендаций о том, как действовать в рамках исследовательской программы. Все науки могут использовать теоретические знания, но это не то же самое, что утверждать, будто все обладающие теорией науки используют ее для направления и организации исследований.
Откуда берется мысль, что в биологии развития именно проблемы задают организационную архитектуру эпистемологии? Дело в том, что они являются распространенной особенностью методов рассуждений в этой области, проиллюстрированной в учебниках. Учебники же фиксируют сложившийся в сообществе консенсус относительно стандартов объяснения, экспериментальных методов, основных понятий и эмпирического содержания. В отличие от учебников по эволюционной биологии, в которых обсуждаются теории естественного отбора, или учебников по экономике, в которых говорится о микроэкономической теории, анализ нескольких изданий основных учебников показывает, что в биологии развития подобных теорий нет.
Книга Основоположения биологии развития Джонатана Слэка (Essential Developmental Biology Slack 2006, 2013) организована вокруг четырех основных типов процессов, которые также описываются как объединенные группы проблем, возникающие во время эмбрионального развития: спецификация областей (формирование паттернов), клеточная дифференциация, морфогенез и рост. Затем эти широкие группы объединяются по стандартной шкале раннего развития, где выделяются гаметогенез, оплодотворение, расщепление, гаструляция и осевая спецификация (см. Раздел 1.2). Различные экспериментальные подходы (клеточная и молекулярная биология, генетика развития и экспериментальная эмбриология) используются на определенном наборе модельных организмов (см. далее раздел 4) для анализа протекания этих феноменов развития. Последующие главы охватывают более поздние стадии развития (например, органогенез), включая различные системы, тщательно рассмотренные на уровнях тканевого слоя, дифференциации и роста, или рассмотренные в отношении эволюционных вопросов (см. далее раздел 5). На протяжении всего этого изложения ни одна конкретная теория, множество гипотез или доминирующая модель не используются для организации столь различных областей исследования. Вместо этого исследовательскую повестку задают широкие группы вопросов, отражающие в целом известные и описанные процессы (дифференциация, спецификация, морфогенез и рост).
Биология развития (Developmental Biology) Скотта Гилберта демонстрирует схожую структуру (Gilbert 2000 [2003, 2006, 2010]). Биологию развития определяют два широких вопроса («Как оплодотворенная яйцеклетка становится взрослым телом? И как это взрослое тело производит еще одно тело?»), которые затем можно подразделить на другие категории: дифференциация, морфогенез, рост, воспроизводство, регенерация, эволюция и регулирование окружающей среды. Эти вопросы еще более аналитически можно раздробить на части при помощи пяти переменных: абстрагирование, разнообразие, связность, положение во времени и пространственный состав. Придаваемые этим переменным значения структурируют комплекс исследовательских вопросов, фигурирующих внутри широкого круга проблем, которые соответствуют вчерне обрисованным нами процессам. Например, исследовательские вопросы относительно событий гаструляции рыбок данио структурированы иначе, чем исследовательские вопросы относительно миграции клеток нервного гребня позвоночных, т.к. включают в себя разные значения пяти упомянутых переменных. Абстрагирование (рыбки данио vs позвоночные), положение во времени (раньше vs позже), пространственный состав (взаимодействие тканевого слоя vs выделенная популяция клеток), разнообразие (эпиболия vs эпителия до мезенхимного перехода) и связность (образование кишечника и энтодерма vs органогенез и эктодерма / мезодерма). Эти конфигурации можно легко модифицировать в ответ на смещения значений различных переменных (Love 2014).[ 6 ]
Этот тщательный анализ проблем – причем с явной эпистемологической структурой, выведенной из разных значений для упомянутых переменных, – сам участвует в организации науки о развитии. Исследователи из разных дисциплин могут работать над одной и той же проблемой, однако ставить разные вопросы, требующие четких и, в то же время, дополнительных методологических ресурсов. Знания и исследования в области биологии развития неразрывно переплетены, только для этого не нужны центральная теория или группа моделей; подобная проблематическая организация эпистемологически доступна всем вовлеченным ученым. Да, теоретические знания, особенно взятые из молекулярно-биологических механизмов (см. molecular biology), или математические модели (например, модели реакции-диффузии), распространены повсеместно (теоретико-информированность). При этом группы проблем, снова и снова возникающие в учебниках и соответствующие различным типам процессов, исполняют роль направляющей архитектуры (нет теоретико-ориентированности): последнюю можно точно описать с помощью приведенных переменных. Возможен и дальнейший анализ этой структуры проблемы, в том числе анализ того, как она изображается в регулярно публикующихся научных статьях, а не только в учебниках, или же в других областях биологии (Brigandt and Love 2012, Love 2008a, 2014).
Объяснительные подходы к развитию
Объяснения в биологии развития обычно являются каузальными, хотя, в отличие от стандартных механистических объяснений, здесь постоянно приобретаются новые способности каузации (в смысле конституирующих сущностей, деятельностей и их организации) посредством развития (Parkkinen 2014). Несмотря на значительный объем проделанной работы по описанию различных аспектов объяснения в биологии развития, совершенно очевидно, что ключевым элементом ее теоретизирования является создание различий, или манипулятивная концепция причинности (Woodward 2003; Strevens 2009; Waters 2007a). Генетические объяснения развития (раздел 3.1), аналогичные тем, которые дает молекулярная генетика, построены на выявлении изменений в экспрессии генов и взаимодействий между их РНК и белковыми продуктами. Эти процессы приводят к изменениям свойств морфологических признаков в онтогенезе (например, формы или размера), в то время как многие другие контекстуальные переменные остаются неизменными. В последнее время растет интерес к физическим объяснениям развития (раздел 3.2), которые обращаются к чисто механическим силам из-за геометрического расположения мезомасштабных материалов, например, потока жидкости (Forgacs and Newman 2005). Исследователи согласны относительно того, какие именно феномены нуждаются в объяснении (раздел 1.2 и раздел 2.2), но расходятся в том, какие факторы имеют бо́льшую объяснительную силу – физические или генетические (Keller 2002).[ 7 ] Существование двух разных типов каузальных объяснений феноменов развития ставит дополнительный вопрос о том, как их можно (если можно вообще) объединить в более целостную объяснительную структуру (раздел 3.3).
Генетика
За последние два десятилетия многие философы обращались к объяснениям развития, чтобы удостоверить или отбросить утверждения о каузальной силе генов (Keller 2002, Neumann-Held and Rehmann-Sutter 2006, Rosenberg 2006, Robert 2004, Waters 2007a). [ 8 ] Генетические объяснения затрагивают философскую тему редукционизма, и, по-видимому, именно они в основном ответственны за эмпирические успехи, накопленные в области биологии развития за последние несколько десятилетий. [ 9 ] Заявления биологов развития зачастую лишь усиливают эту точку зрения:
Биология развития ... имеет дело с процессом, посредством которого гены в оплодотворенной яйцеклетке контролируют поведение клеток в эмбрионе и тем самым определяют его паттерны, его форму и большую часть его поведения ... дифференциальная активность генов контролирует развитие. (Wolpert et al., 1998: v, 15)
Утверждения такого типа усиливаются в обращениях к генетической программе развития.
[Элементы генома] содержат секвенционально-специфицированный код развития. Они определяют конкретный результат процессов развития и, следовательно, форму животного, развившегося из каждого эмбриона. ... Развитие – это выполнение генетической программы по построению организма данного вида (Davidson 2006: 2, 16). [ 10 ]
Обоснованность этих утверждений была предметом напряженных дискуссий. [ 11 ] Самые сильные утверждения о генетических программах или генетическом контроле развития имеют эмпирические и концептуальные недостатки: невнимание к пластичности и роли окружающей среды, двусмысленность в отношении местоположения действующей агентности и зависимость от метафор, взятых из компьютерных наук (Gilber and Epel 2009, Keller 2002, Moss 2002, Robert 2004). Но эти контраргументы не затрагивают имеющийся в молекулярной генетике принцип генетического объяснения, заключающийся в производстве изменений (difference-making principle) (Waters 2007a). Последний производит более узкие и точечные каузальные утверждения [т.е. утверждения формы «А было причиной Б» – прим. ред.] в контролируемых экспериментальных условиях и применим к различным молекулярным образованиям, которые играют каузальную роль во время развития. Например, к регуляторным РНК, белкам и сигналам из окружающей среды. Мы можем это увидеть, вновь обратившись к примеру кардиогенеза позвоночных (раздел 1.2).
Проблематично ли утверждение, что гены содержат всю информацию, необходимую для формирования сердца позвоночных? Существует ли генетическая программа в ДНК, контролирующая развитие сердца? Являются ли гены основным поставщиком и организатором материальных ресурсов для развития сердца, во многом определяющим фенотипический результат? Проводившиеся исследования развития сердца установили роль гидродинамических сил в спецификации внутренней формы сердца (Hove et al., 2003) и ее право-/левой асимметрии (Nonaka et al., 2002). Биохимические градиенты внеклеточного кальция ответственны за активацию асимметричной экспрессии регуляторного гена Nodal (Raya et al., 2004), тогда как ингибирование градиентов напряжения скремблирует нормальную асимметрию (Levin et al., 2002). Некоторые гены определенно являются факторами влияния в этих процессах (Srivastava 2006, Brand 2003, Olson 2006), но вывод, что гены содержат всю информацию, необходимую для создания особенностей формы сердца, необязателен и не обоснован. Хотя в некоторых случаях можно гарантировать, что различия в ДНК-последовательностях являются каузальными факторами в конкретных процессах онтогенеза (Waters 2007a), – например, иерархически организованные сети генетических факторов влияния, объясняющие спецификацию тканей (Peter and Davidson 2011), – тем не менее разнообразие сущностей, к которым обращается в молекулярная генетика, степень их индивидуальной и совместной роли в определении результатов развития означают, что дискуссии о значении, сфере охвата и силе генетических объяснений будут продолжены (Griffiths and Stotz 2013). Но переход от генетических программ и генетического детерминизма к ДНК, РНК и белкам как к действующим совместно факторам влияния предполагает, что мы концептуализируем и другие причинные факторы сходным образом.
Физика
Поток жидкости, рассмотренный как физическая сила, также оказывает значительное влияние на процесс развития сердца и на онтогенез в более общем плане. Биологи развития обращаются к подобным физическим источникам влияния, понимая их как факторы, проявляющие себя в морфологических свойствах явлениях развития (Forgacs and Newman 2005). Физический причинно-следственный подход начал преобладать в первой половине XX в. благодаря работе Д'арка Томпсона (Thompson 1992 [1942], Keller 2002: ch. 2, Olby 1986). Это происходило на фоне повышенного внимания к хромосомной теории наследования и попыток изучить явления развития при помощи классических генетических методов (Morgan 1923, 1926, 1934). Томпсон обратился к различиям в темпах роста и к ограничениям геометрических соотношений [частей организма], чтобы объяснить, из чего возникает конкретная морфология организма. Визуальные репрезентации абиотических, механических аналогов обеспечивали определенное правдоподобие: например, удачно объяснялась форма жидких брызг или висячих капель в случае чашевидных и колоколообразных конфигураций свободноплавающей медузы в половой стадии. Если физические силы создают характерные морфологии в вязкоупругих материалах, тогда аналогичные морфологии, обнаруживаемые у живых существ, следует объяснять с точки зрения физических сил, действующих на вязкоупругие материалы развивающегося эмбриона. Однако морфогенетические процессы, благодаря которым складываются форма и структура морфологии, за последние полвека рассматривались прежде всего, – если не исключительно, – с точки зрения генетики. Физические подходы сместились на периферию, в то время как подходы, предлагаемые молекулярной генетикой, стали все более влиятельными (Fraser and Harland 2000).
«Молекуляризация» экспериментальной эмбриологии является одной из самых ярких историй успеха в современной биологии, поскольку было обнаружено, что гены и их взаимодействия (например, в транскрипционных сетях и сигнальных путях) лежат в основе конкретных особенностей дифференциации, морфогенеза, формирования паттернов и роста в период возникновения структуры в ходе развития. В современной биологии развития преобладают генетические подходы; физические типы причинности, как правило, игнорируются или же рассматриваются как незначительные. Среди исследователей, интересующихся проявлениями физической причинности во время эмбриогенеза, стало чувствоваться разочарование.
Для молекулярных типов объяснения причиной выступает молекула или ген. Объяснить феномен означает, по сути, идентифицировать гены и охарактеризовать белки, без которых он перестанет существовать или окажется за пределами нормы. Молекула служит объяснением, [физическая] сила – описанием; если вы станете утверждать нечто иное, это в лучшем случае вызовет сожаление (Albert Harris to John Trinkaus, 12 March 1996; Источник: Архивы библиотеки морской биологической лаборатории).
Несмотря на подобное преобладание генетических подходов к объяснению и разочарование среди исследователей, использующих другие подходы, вокруг физических объяснений развития видны признаки надвигающегося массового энтузиазма, причем особый интерес вызывает возможность их объединения с генетическими объяснениями (Miller and Davidson 2013).
Объединение подходов: генетика и физика
Томпсон считал, что физические силы имеют объяснительный потенциал, но самих по себе их недостаточно для описания того, как в ходе развития возникает морфология; наследственность (генетика) также была необходимым каузальным фактором. [ 12 ] Тем не менее Томпсон довольно быстро подметил, что в разгар растущего внимания к наследственности (генетике) механические типы причинности могут уйти в забвение:
будет по меньшей мере преувеличением, если мы решим пренебречь этими прямыми физическими и механическими типами причинности и увидим в особенностях кости́ исключительно результат вариации и наследственности. (Thompson 1992 [1942]: 1023)
Несмотря на подобное преувеличение, возникавшее на протяжении большей части XX века, задача объединения – или интеграции – этих двух подходов теперь стала очевидна. [ 13 ]
Ни у кого нет сомнений насчет того, действуют ли генетические и физические типы причинности одновременно:
как физические факторы, так и биохимические сигнальные пути эмбриона вносят вклад в порождение формы организма. (Von Dassow et al. 2010: 1)
То есть они не являются конкурирующими объяснениями одного и того же феномена. Наоборот, объяснения должны учитывать, что взаимовлияния этих причин продуктивно воздействуют на процесс развития:
все большее число примеров указывает на существование двустороннего взаимодействия между экспрессией некоторых генов, играющих важную роль в развитии, и механическими силами, связанных с морфогенетическими движениями. (Brouzés and Farge 2004: 372)
Генетические причины могут привести к физической каузальности и наоборот. Физическая причинность приводит к генетической каузальности посредством механотрансдукции. Растяжение, противодействие, сжатие, напряжение сдвига потока и другие проявления физической динамики ощущаются различными молекулярными компонентами внутри и снаружи клеток, которые затем переводят эти изменения окружающей среды в биохимические сигналы (Hoffman et al., 2011; Wozniak and Chen 2009). Генетическая каузальность приводит к физической каузальности путем порождения различных физических свойств у клеток и тканей, в частности, за счет присутствия, отсутствия или изменения частотности конкретных белков. Так, различные паттерны экспрессии молекул клеточной адгезии (например, кадгеринов) могут приводить к различиям в адгезии вдоль эпителиальных листов ткани, и, тем самым, создавать разделение фаз или ячейки через изменение поверхностного натяжения (Newman and Bhat 2008). Если эти типы причинности не конкурируют друг с другом, то как тогда можно объединить генетические и физические факторы влияния в комплексное причинное объяснение? Какое может объяснительное единство быть достигнуто для этих «двусторонних взаимодействий»?
Поиск философских моделей для интеграции объяснений генетики и физики остается открытой проблемой (cv/ предстоящую публикацию Love). Распределение каузальной ответственности – в смысле определения того, какой вклад вносит та или иная причина (например, совокупности каузальных величин между различными физическими силами в механике Ньютона), проблематично, поскольку для этого требуется, чтобы производимые ими следствия были соизмеримы (Sober 1988). В контексте причинности, понимаемой с точки зрения факторов влияния, трудность интеграции объяснений – это, по сути, разновидность проблемы каузального мышления, обнаруженной Джоном Стюартом Миллем и названной им «смешением следствий»: в этой ситуации имеют место несколько причин, смешанным образом вносящие вклад в получение результата.
Это затруднение особенно заметно в случае физиологических явлений; редко когда удается разделить между собой различные действующие силы, в совокупности составляющие организованное тело, не уничтожая вместе с тем исследуемые нами феномены. (Mill 1843 [1974]: 456 [book 3, chapter 11, section 1, paragraph 7])
Аккуратное использование статистической методологии в экспериментах может ответить на вопрос, ответственен ли один из факторов влияния за большее (чем другой) влияние на вариацию следствий для конкретной популяции. Однако ранжирование каузальных факторов в отношении произведенных ими изменений – отнюдь не то же самое, что объединение двух типов каузальности в одно общее описание. Другой подход к проблеме интеграции заключается в сведении всех каузальных взаимодействий только к одному из двух типов, благодаря чему достигается своеобразное объяснительное единство (Rosenberg 2006). Однако работающие в этой области биологи отказываются от такого подхода, полагая, что и генетический, и физический тип каузальности по-своему значимы и не сводимы друг к другу.
Другая стратегия носит название интегративного плюрализма (Mitchell 2002). Этот подход включает в себя двухэтапную процедуру объяснения сложных явлений, чьи особенности формируются под воздействием нескольких причин: этап (а) формулировка идеализированных моделей, где отдельные причинно-следственные факторы действуют изолированно («теоретическое моделирование»); этап (б) объединение идеализированных моделей для объяснения того, как определенные, конкретные явления возникают из комбинации данных причин. Эта модель наводит на размышления, но также имеет существенные недостатки: так, в биологии развития каузальное мышление не предполагает формулировки идеализированных моделей, поэтому предполагаемая интеграция лишается требуемой определенности. Еще одну возможность дает интеграция генетических и физических факторов влияния в один механизм (Darden 2006; Craver 2007). Хотя ценность этого варианта состоит в том, что в нем подчеркивается продуктивная преемственность между факторами влияния на разных этапах последовательности (то есть последовательности их двусторонних взаимодействий), он также имеет недостатки. К ним относятся:
Разнонаправленность подходов к измерению времени. Вместо времени «в механизме», измерение времени производится внешними стандартизованными стадиями (см. далее, Раздел 5.2). Стадии облегчают изучение разнообразных механизмов развития, с различными характерными скоростями и продолжительностью для соответствующих стадий в рамках общей структуры данного модельного организма (например, Drosophila). Кроме того, с помощью стадий можно изучать консервативные молекулярные механизмы у разных видов, поскольку соответствующее описание механизма не привязано ко временной последовательности модельного организма.
Ожидание того, что описание механизмов «достигнет дна», т.е. молекулярных образований самого низкого уровня активности (Darden, 2006). В случае объединения генетических и физических факторов влияния двухстороннее взаимодействие означает, в том или ином типе каузальности «достижение дна» старательно избегается.
Требование стабильной, составной организации для механизмов:
Механические объяснения – это конститутивные объяснения или объяснения через компоненты: они объясняют поведение механизма в целом с точки зрения организованной деятельности и взаимодействия ее компонентов. (Craver 2007: 128)
Однако эти описания механизмов часто встроены в различные контексты развития (происходящие в разное время в онтогенезе) с различными композиционными отношениями (внутри вида и между видами). Двухстороннее взаимодействие между генетическими и физическими факторами влияния не поддерживается – в данной объяснительной модели – именно потому, что упомянутые композиционные различия изменяют отношения физической причинности (поток жидкости, напряжение и т.д.). Биологи развития смогли довольно широко обобщить отношения генетической причинности (в терминах генетических механизмов) по видам, но попытка объединить их с физической причинностью потребовала сужения сферы каузальных утверждений.
Адекватные философские модели систематической зависимости между генетическими и физическими факторами влияния в онтогенезе должны учитывать, как временные отношения, необходимые для создания каузальных связей, укоренены во внешней периодизации, используемой биологами развития. Наложение разных временных масштабов способно привести к тому, что различные факторы приобретут значимость или выраженность, что важно для установления того, как различные типы причин могут быть объединены в интегрированные объяснения. Одна из возможностей заключается в сопоставлении этих факторов влияния на разных этапах с помощью экспериментальной проверки – таким образом, чтобы они продемонстрировали производящую непрерывность [своих следствий] в рамках, установленных внешней периодизацией (см. предстоящую публикацию Love). Такой подход облегчает отображение симметрии между факторами влияния, поскольку генетические факторы влияния могут быть размещены до или после физических (и наоборот). Хотя это и не дает нам способа, с помощью которого мы могли бы складывать каузальные величины (как векторное сложение в Ньютоновской механике), описанный подход предлагает внятную стратегию распределения ответственности между различными причинами при помощи временно́й организации, что выходит за рамки простого ранжирования факторов влияния. Периодизация выступает шаблоном из практики биологов развития, который обеспечивает целостность или единство различным типам причинности, и призван дать хотя некое интегрированное объяснение морфологии, возникающей в результате последовательности процессов развития.
Не все типы каузальных объяснений включают внешнюю периодизацию: существуют и другие способы объединения причин, призванные создать более интегрированную объяснительную рамку. Одна область, в которой анализируются обобщенные объяснения феноменов развития, относится к описанию механизмов и математическому моделированию в системной биологии (Brigandt 2013, Fagan 2013). Например, Фаган (Fagan 2013: ch. 9) показывает, как интегрированное объяснение возникает из пошаговой процедуры – она начинается с подробного описания молекулярного механизма, за которым следует формулировка абстрагированной схемы соединений (wiring diagram) взаимодействий между компонентами, которая затем переводится в систему уравнений, способную с течением времени учитывать изменения в межкомпонентных взаимодействиях. Решения этих систем уравнений и наложение решений для взаимодействий компонентов системы на поведение всей системы в рамках общего панорамного представления дает более систематическое объяснение клеточной дифференциации.
Модельные организмы для изучения развития
Модельные организмы играют центральную роль в современной биологии и исследованиях эмбриогенеза (Ankeny and Leonelli 2011, Steel 2008, Bier and McGinnis 2003, Davis 2004). Биологи используют лишь небольшое количество видов для экспериментального установления различных свойств онтогенеза (например, C. elegans , Drosophila и Brachydanio [рыбка данио], см. Рисунок 4). Эти экспериментальные модели позволяют исследователям действительно глубоко изучить развитие и упростить раскрытие каузальных связей. Критики сомневаются в том, хорошо ли эти модели представляют другие виды из-за присущих им систематических отклонений – таких как быстрое развитие и короткое время генерации (Bolker, 1995), а также из-за проблематических предположений о консервативности генных функций и регуляторных сетей (Lynch 2009). Например, эмбриогенез C. elegans не является репрезентативным для нематод с точки зрения формирования рисунка и спецификации клеток (Schulze and Schierenberg 2011), а формирование придатков у рыбок данио – плохой образец развития придатков у тетраподов (Metscher and Ahlberg 1999).

Рисунок 4: Drosophila melanogaster (дрозофила обыкновенная) является одним из стандартных модельных организмов, используемых в биологии развития.
Один из ответов на эту критику заключается в подчеркивании консервативных генетических механизмов, общих у всех животным, несмотря на различия в конкретных феноменах развития (Gerhart and Kirschner 2007, Ankeny and Leonelli 2011, Weber 2005). Фруктовые мухи могут быть нерепрезентативными в плане проявления синцитиального развития, но они используют коллинеарную экспрессию генов Hox для разметки осей своего тела. Данный ответ указывает на то, что репрезентативность или нерепрезентативность модельного организма самого по себе – слишком грубый критерий для определения целесообразности его использования. В отношении модельного организма мы должны спросить, что именно он репрезентирует, и некоторые последовали именно в этом направлении. Джессика Боллер различала образцовые и суррогатные формы представления (Bolker 2009): первые служат фундаментальным исследованиям, выступая образцами для большей группы, тогда как последние соответствуют моделям, предназначенным для обеспечения косвенного экспериментального доступа к другим недоступным феноменам, в частности, таким как мышиные модели человеческих психологических расстройств (например, шизофрении). Суррогатные модели принимаются в области биомедицины, где представляющие интерес феномены проявляются у людей. Большинство биологов развития рассматривают модельные организмы как образцы, нежели как суррогаты. [ 15 ] Таким образом, чтобы ответить на критику о нерепрезентативности, критерий представительства должен быть рассмотрен более подробно. [ 16 ]
Основное допущение относительно модельных организмов заключается в том, что они разделяют соответствующие отношения подобия с более крупными группами животных. Это допущение является примером ответа вопрос о том, что значит для что для моделей в науке (см. models in science) репрезентировать некий феномен. Модельные организмы представляют феномены развития у видов, которые либо мало изучены, либо вообще не изучаются: «мы изучаем мух и лягушек в качестве примеров развития животных в целом» (Nüsslein-Volhard, 2006: 87). Одна из причин уверенности в том, что они действительно являются образцами, проистекает из индуктивного вывода из обнаруженных паттернов эволюционной консервативности в отношении феноменов развития (например, гаструляции или образования сомитов). Если все или большинство модельных организмов обладают некой особенностью развития, тогда все или большинство животных также будут иметь эту особенность. Данный вывод можно более или менее узко ограничить (например, если все или большинство позвоночных модельных организмов образуют сомиты, тогда все или большинство позвоночных видов также будут иметь эту особенность).
Благодаря такой степени достоверности модельный организм («источник») может представлять другие, неизученные виды («мишени» [«targets»]). Приведенное базовое различие между моделью или источником – и феноменами или мишенями, которые предполагается представить, – повсеместно встречается в рассуждениях о модельных организмах (Ankeny and Leonelli 2011). Рыбка данио – это модель или репрезентация развития позвоночных, целевые (target) феномены, т.к. мы ожидаем узнать о развитии позвоночных вообще, изучая протекание онтогенеза конкретно у данио. Мы не тратим время и ресурсы на рыбу данио в качестве модельного организма потому лишь, что нас интересуют рыбы данио сами по себе. Мы планируем, исходя из наблюдений за рыбками данио, сделать утверждения о формировании сомитов: эти утверждения будут применяться к формированию сомитов у других позвоночных, на исследование которых у нас никогда не найдется времени или денег.
Биологи развития часто говорят об исследованиях механизмов, ответственных за феномены онтогенеза, и сосредотачиваются на консервативных генетических и клеточных механизмах в модельных организмах (Gerhart and Kirschner 2007, Ankeny and Leonelli 2011, Weber 2005). Это предполагает различие между репрезентацией феноменов развития и репрезентацией генетических и клеточных механизмов, участвующих в процессе развития. Если мы стремимся объяснить развитие сердца (феномены), мы можем исследовать молекулярные или клеточные механизмы, действующие в области сердца во время онтогенеза у рыбок данио. Некоторые из этих механизмов могут оказаться консервативными, даже если феномены развития таковыми не являются. У мухи Drosophila есть только один тип сердечных клеток, у нее нет клеток нервного гребня, а морфология ее сердца не имеет предсердной или желудочковой камеры (Kirby 1999). Однако у всех исследованных до сих пор беспозвоночных и позвоночных кардиогенез существенно зависит от экспрессии гомеобокса гена Nkx2-5 / tinman (Gajewski and Schulz, 2002). Допустима также и обратная ситуация: могут наблюдаться схожие феномены развития, но производящие их генетические и клеточные механизмы могут различаться. У амфибий нервная трубка (нейруляция) образуется в ходе процесса инвагинации (сгибания эпителиального листа), тогда как у костистых рыб нервная трубка образуется путем кавитации (исчезновения массива ткани из-за гибели клеток). Нейронная трубка гомологична у позвоночных (т.е. феномен консервативен), но участвующие в инвагинации и кавитации клеточные и генетические механизмы различны (Davies 2005: ch. 4).
Различие между явлениями (феноменами) и механизмами предполагает специфичность; то есть, если судить о зависимости между источником (моделью) и мишенью (target), существуют специфические явления (образование сомитов у позвоночных) или механизмы (коллинеарная экспрессия гена Hox). Однако развитие животных складывается из множества различных процессов, включающих в себя множество различных механизмов. Вот почему еще одно различие, важное для критерия репрезентативности, относится к вопросам специфичности vs. разнообразия при выборе и использовании модельных организмов. Модель может представлять собой один тип целевые (target) феноменов (дифференциация или рост) или механизм (сигнальная система клетки или клеточный цикл), но не других, – это специфичность, – или может делать это лучше или хуже по отношению к определенным типам явлений или механизмов. С другой стороны, модель может представлять собой несколько типов целевых феноменов и механизмов одновременно, – это разнообразие, – при этом каждый из этих типов может быть представлен различным образом. Существуют определенные компромиссы по поводу того, насколько хорошо различные явления или механизмы могут быть совместно представлены в модельном организме. Отметим, что экспериментальные организмы могут быть выбраны по критериям разнообразия и специфичности одновременно: например, когда биолог, работающий над конкретным феноменом, намеревается работать над другими, используя тот же модельный организм в будущем. Также выбор может быть сделан в пользу только одного из этих двух аспектов. Модель может быть подходящей, если она может репрезентировать как механизмы, так и явления, даже не будучи наилучшим представителем каждого конкретного механизма или явления. И наоборот, модельный организм может быть подходящим, если он является наилучшим представителем конкретного механизма, несмотря на то, что является крайне плохой моделью для других феноменов или механизмов. Разнообразие свидетельствует о том, что моделью является «весь организм».[ 17 ] Можно указать на еще одно различие – различие между «модельными организмами» и «экспериментальными организмами» (Ankeny and Leonelli 2011) или «общими модельными организмами» и «модельными организмами по принципу Крога» (Love 2010a). При отборе и использовании общих модельных организмов ученые преимущественно руководствуются критерием репрезентативности; при использовании и отборе экспериментальных или модельных организмов по принципу Крога руководствуются критерием специфичности.
Другие относящиеся к критерию репрезентации вопросы включают в себя: как использование модельных организмов становится примером рассуждений на основе прецедентов (Ankeny 2012), и как использование модельных организмов влечет за собой идеализации неких черт или известные отклонения от этих черт, присутствующих в мишени (target) данной модели как результат лабораторного культивирования (Ankeny 2009). Подготовка модельных организмов для экспериментального анализа предполагает преднамеренное искажение механизмов или феноменов, например, как в случае стандартизированной периодизации онтогенеза, искажающей степень и виды изменений в процессе развития (Love 2010a; Раздел 5.2). Кроме того, вопрос о репрезентации – далеко не единственный из тех, которые было бы уместно задать относительно того, как мы понимаем использованием модельного организма. Поскольку модельные организмы используются для экспериментального вмешательства, вопросы репрезентации должны быть сопоставлены с вопросами манипуляции (см. Дополнение к данному разделу: Модельные организмы и манипуляция).
Развитие и эволюция
Отношения, складывающиеся между развитием и эволюцией, сложны и являются предметом все новых исследований (см. обзор в: Love 2015). В нестройном конгломерате исследовательских программ доминируют два основных направления (Raff 2000, Müller 2007): (a) эволюция развития, или исследование паттернов и процессов изменений, протекающих в онтогенезе с течением времени; и (б) изучение процессов развития, лежащих в основе эволюции, или исследование каузального воздействия онтогенетических процессов на эволюционные траектории (как с точки зрения возникновения ограничений, так с точки зрения того, как они им содействуют). В данном разделе рассматриваются два примера столкновений концепций и практик биологии развития и эволюционной биологии: проблематичное обращение к функциональной гомологии в генетике развития, которое призвано подкрепить эволюционные обобщения относительно онтогенеза (раздел 5.1); а также – напряжение между использованием нормальных стадий для исследования развития и определением эволюционной значимости фенотипической пластичности ( раздел 5.2 ). Эти случаи раскрывают некоторые философские проблемы, присущие вопросу о том, как возможно
связать развитие и эволюцию.
Функциональная гомология в генетике развития.
Консервативная роль генов Hox в формировании осевых паттернов (axial patterning) у животных называется функционально гомологичной (Manak and Scott 1994), в дополнение к отношению структурной гомологии, устанавливающегося между последовательностями ДНК. И все же «функциональная гомология» является противоречием в терминах (Abouheif et al., 1997), поскольку по определению гомолог это – «один и тот же орган у разных животных при любом разнообразии формы и функции» (Owen 1843: 379) – потомок; эволюционное различие между гомологией (структурой) и аналогией (функцией) основано на признании этого обстоятельства. Следовательно, идея функциональной гомологии оказывается теоретически путанной, и в ее использовании молекулярными биологами развития существует концептуальная напряженность.
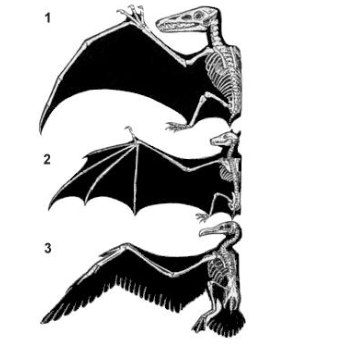
Рисунок 5: Крылья позвоночных, как передние конечности, гомологичны; они имеют одно и то же происхождение от одной и той же структуры. Функция крыльев у позвоночных (т.е. полет) аналогична; хотя крылья выполняют сходные функции, их роль в полете эволюционировала раздельно.
Ссылка на «орган» в определении Оуэна указывает на структуру (сущность), найденную в организме, которая может отличаться по своим очертаниям и составу (форме) или своей функцией у видов, у которых она встречается. В контексте эволюции это будет означать, что тождественность оплачивается ссылкой на происхождение от общего предка. Так как структуры могут быть схожи и в силу естественного отбора, действующего в похожих средах, гомология противопоставляется аналогии. Гомологичные структуры одинаковы в силу происхождения от общего предка, независимо от того, в исполнение каких функций эти структуры участвуют, тогда как аналогичные структуры схожи в силу процессов отбора, благоприятствующих сопоставимым функциональным результатам, вне зависимости от общности происхождения (рис.5).
Именно это делает сходство функции особенно проблематичным критерием гомологии (Abouheif et al., 1997). Поскольку функциональное сходство и так является надлежащей связью для аналогии, аналогам не обязательно иметь такую же функцию как следствие происхождения от общего предка – несмотря на различное происхождение, схожести оказывается достаточно (Ghiselin 2005). В число классических случаев аналогии входят таксоны, не имеющие друг с другом недавнего общего предка, имеющего такую же структуру. Пример такого таксона – внешняя морфология тела дельфина и тунца (Pabst 2000). Таким образом, функциональная гомология, похоже, является категориальной ошибкой, поскольку то, что делает структура, не должно приниматься в расчет при оценки соответствия гомолога; сходство же функции часто является результатом адаптации путем естественного отбора к общим требованиям окружающей среды, нежели проистекает из общей родословной.
Хотя мы можем быть склонны просто запретить терминологию функциональной гомологии, ее широкое использование в молекулярной биологии и биологии развития должно по крайней мере заставить нас задуматься: «Функциональная гомология фосфатаз человека и делящихся дрожжей Cdc14» (Vázquez-Novelle et al., 2005). [ 18 ] Хотя и важно признать повсеместность этой практики, некоторые случаи все же могут быть незаконными. Замена структурно гомологичных генов между различными видами для спасения мутантных или «нулевых» фенотипов не является подлинным критерием функциональной гомологии, особенно когда установлению филогенетического контекста либо уделено мало внимания, либо не было уделено вовсе. Это делает многие утверждения о функциональной гомологии между модельными организмами как минимум подозрительными. Чтобы избежать концептуальных трудностей, нужно уделять пристальное внимание значению «функции». Биологическая практика дает примеры использования функции по меньшей мере в 4-х различных значениях (Wouters 2003, 2005): деятельность (то, что нечто делает), каузальная роль (вклад в способность), пригодность или жизнеспособность (ценность наличия чего-либо), а также селекционный эффект или этиология (возникновение и поддержание посредством естественного отбора). Споры разгорелись по поводу того, какое из значений лучше всего подходит (если подходит вообще) для различных аспектов биологических и психологических рассуждений, или же является наиболее общим по своему охвату (т.е., что делает их все понятиями «функции»?); как бы то ни было, в нашем случае единственный вопрос состоит в том, можем ли мы определить правомерное понятие гомологии функции.
Если мы не хотим смешивать гомологию и аналогию, то наше понимание функции не может основываться на истории отбора, связанной с понятием аналогии и касающейся определенного разнообразия функции. Точно так же интерпретации, которые основываются на критерии жизнеспособности, концентрируются на особенностях, в которых разнообразие функций имеет решающее значение, поскольку дает бо́льшие шансы на выживание. Любая интерпретация понятия функции, опирающаяся на конкретную вариативность функции (потому, что она была отобрана, или потому, что она обеспечивает жизнеспособность) сталкивается с требованием, чтобы гомология относилась к чему-то «при любом разнообразии формы и функции». Интерпретация функции через ее каузальную роль подчеркивает системную способность, в которую функция вносит свой вклад. Она также фокусируется на определенном разнообразии функции, хотя и иначе, чем в интерпретациях селекционного эффекта или жизнеспособности. Только интерпретация активности («то, что нечто делает») делает акцент на самой функции, помимо ее специфического вклада в системную способность и положения в более широком контексте. Следовательно, «активность-функция» является наиболее подходящим значением для понятия «гомологии функции», поскольку по крайней мере возможно, активность-функции будут оставаться постоянными при любом разнообразии. Оценка тождественности, обусловленной общей родословной, производится отдельно от оценки функции (или ее использования), понимаемой в терминах казуальной роли, преимуществ в выживании или истории отбора. [19] Активность-функции могут использоваться по-разному, имея при этом общее происхождение предка (т.е. являясь гомологичными). Точнее, гомология функции может быть определена как одна и та же активность-функция у разных животных при любом разнообразии формы и функции-использования (Love 2007). Этот вариант однозначно устраняет напряжение, которым страдает понятие функциональной гомологии.
Тщательное обсуждение регуляторной функции генов в развитии и эволюции позволяет распознать нечто похожее на различие между активность-функцией и функцией-использования (т.е. между тем, что ген делает, и тем, для чего он нужен в некотором процессе организма).
При изучении молекулярной эволюции регуляторных генов, их биохимическая функция и функция развития должны рассматриваться раздельно. Биохимическая функция PAX-6 и незрячесть выступают общими факторами транскрипции (которые связывают и активируют нижеследующие гены), но их функция развития — это их специфическое участие в морфогенезе глаза (Abouheif 1997: 407).
Биохимическая функция – это активность-функция, а функция развития – это функция-использование. Данное различие помогает провести границы между расходящимися эволюционными траекториями. Биохимические (активность-функции) генов часто консервативны (т.е. гомологичны), будучи одновременно доступны для кооптации для выполнения каузальной роли (использование-функция) в отдельных процессы развития. Те же регуляторные гены эволюционно устойчивы с точки зрения активность-функции и эволюционно лабильны с точки зрения функции-использования. [20] Подразумевается, что утверждения о гомологии функций-использования для генов как функции развития являются подозрительными по сравнению с утверждениями о гомологии активность-функций для генов как биохимических функций, т.к. функции развития, вероятнее всего, изменились по мере увеличения филогенетического расстояния.
Различие между биохимической (активность) функцией и функцией развития (использования) подкрепляется иерархическими аспектами гомологии (Hall 1994). Способность, определяющая функцию-использования регуляторного гена на одном уровне организации (например, в формировании осевых направлений), сама должна рассматриваться как активность-функция на другом уровне организации: таком, как дифференциация последовательно повторяющихся элементов вдоль оси тела. (Обратите внимание, что «уровень организации» не обязательно должен быть составным, и, следовательно, язык «высшего» и «нижнего» уровней может быть неуместным.) Роль, которую играют гены Hox в формировании осевых паттернов, может быть консервативной в силу их биохимических гомологий активности-функции, однако гены Hox не являются гомологами функций-использования именно из-за их роли в развитии. Вместо того, чтобы сосредоточиться на активности генетического компонента и его каузальной роли в формировании осевых паттернов, мы переходим к активности самого формирования осевых паттернов и его каузальной роли где-либо (или когда-либо) в эмбриональном развитии.
Правомерная, с концептуальной точки зрения, идея гомологии активность-функции нужна не для того, чтобы упорядочить идеи биологии развития. Она помогает в интерпретации экспериментальных данных и ограничивает сделанные выводы. Например, гены NK-2 участвуют в спецификации мезодермы, а это лежит в основе морфогенеза мышц. У Drosophila экспрессия определенного гена NK-2 (tinman) имеет решающее значение для развития сердечной и висцеральной мезодерм. Если tinman выбивается и трансгенно заменяется своим ортологом позвоночных, Nkx2-5, сохраняется только спецификация висцеральной мезодермы; регуляция сердечной мезодермы не происходит (Ranganayakulu et al., 1998). Область белка у позвоночных около 5'-конца полипептида достаточно сильно отличается, чтобы не допустить соответствующей регуляции морфогенеза сердца. Гомеодомены (участки последовательности, обеспечивающие связывание ДНК) для позвоночных Nkx2-5 и Drosophila tinman взаимозаменяемы. Неспособность Nkx2-5 сохранить спецификацию сердечной мезодермы не связана с активность-функцией различительного связывания ДНК. Один компонент ортологичных (гомологичных) белков у обоих видов сохраняет гомологию активность-функции, связанную с висцеральной мезодермой, но другой компонент (а не гомеодомен) расходится. Этот гомеобокс гена не имеет единственной функции-использования (как ожидалось), но также он не имеет одной активность-функции. Любая адекватная оценка этих случаев должна признавать более тонкоструктурированное разложение генов на рабочие единицы, чтобы ухватить подлинную консервативность активность-функции. Мы можем связать гомологи активность-функций непосредственно со структурными мотивами внутри гена, однако во всей открытой рамке считывания не обязательно существует лишь одна активность-функция.
Снижение концептуальной напряженности между биологией развития и эволюционной биологией в их отношении к гомологии функции напрямую влияет на каузальные обобщения и выводы, получаемые из исследований модельных организмов (раздел 4). Активность-функциональная гомология направляет наше внимание на стабильность или консервативность активности. Подобная консервативность указывает, когда изучение механизмов в модельных организмах приведет к надёжным и стабильным обобщениям. Широкое использование функциональной гомологии в биологии развития направлено именно на такого рода вопрос, объясняющий свою живучесть в экспериментальной биологии, несмотря на концептуальные двусмысленности. Обобщения, касающиеся молекулярных сигнальных каскадов, подтверждены согласованными биохимическими активностями, а не ролями развития (хотя иногда они могут совпадать). Таким образом, детали активность-функции насчёт сигнального каскада, полученные из модельного организма, могут быть обобщены посредством гомологии с другими неизученными организмами, даже если у других видов для активность-функции роль в развития варьируется.
Нормальные стадии и фенотипическая пластичность
Любые теоретические стратегии сочетают в себе отличительные преимущества наряду со скрытыми слабостями. Например, разложение системы на составляющие для понимания демонстрируемых ей особенностей способствует раскрытию каузальных взаимодействий локализованных составляющих, но одновременно с этим преуменьшает взаимодействие с элементами, внешними по отношению к системе (Wimsatt 1980; Bechtel and Richardson 1993). Иногда описательная и объясняющая научная практика успешна именно потому, что науки намеренно игнорируют отдельные аспекты природных явлений или используют различные техники приближения. Идеализация – это одна из стратегий теоретизирования, используемая учеными для описания, моделирования и объяснения; она целенаправленно абстрагируется от особенностей, о существовании которых ученые осведомлены. Например, внутреннее пространство клетки часто изображается как относительно пустое, хотя известно, что внутриклеточное пространство заполнено (Ellis 2001); переменная клеточного объема принимает значение, которое заведомо является ложным (т.е. относительно пустым). Идеализации самым различным образом сознательно игнорируют вариации свойств или исключают конкретные значения для переменных во имя описательных и объяснительных целей (Jones 2005, Weisberg 2007).
Изучение онтогенеза в модельных организмах (Раздел 4) производится с помощью выделения набора нормальных стадий эмбрионального развития. Траектория развития от оплодотворенной зиготы до полностью сформировавшейся взрослой особи распадается на отдельные временные периоды, выделенные с учетом протекания значимых событий, таких как оплодотворение, гаструляция или метаморфоза (Minelli 2003: ch 4; см. Раздел 1.2). Это позволяет исследователям в различных лабораторных условиях стандартизовать процедуру сравнения экспериментальных результатов (Hopwood, 2005, 2007). Такие результаты имеют решающее значение для больших сообществ биологов развития, работающих с хорошо зарекомендовавшими себя моделями, такими как цыпленок (Hamburger and Hamilton, 1951) или рыбка данио (Kimmel et al., 1995): «Сегодня эмбриологические исследования нельзя представить без таких стандартных серий» (Hopwood 2005: 239). Эти нормальные стадии представляют собой форму идеализации, поскольку намеренно игнорируют некоторые виды изменений в развитии, в том числе вариации, связанные с переменными окружающей среды. Хотя это и облегчает изучение конкретных каузальных связей, однако специфические виды изменений в характеристиках развития, возможно, относящиеся к эволюции, сводятся к минимуму по мере того, как онтогенез становится экспериментально управляемым (Love 2010a).
Фенотипическая пластичность является повсеместным биологическим явлением. Она включает в себя способность определенного генотипа производить фенотипические вариации – часто в форме качественно отличных фенотипов – в ответ на перепады сигналов, поступающих из окружающей среды (Pigliucci 2001; DeWitt and Scheiner 2004, Kaplan 2008, Gilbert and Epel 2009). Один из известных примеров – сезонные гусеничные морфы, зависящие от разных источников питания (Greene 1989). В числе релевантных переменных окружающей среды есть также: температура, питание, давление/гравитация, свет, хищники или стрессовые условия, плотность популяции (Gilbert and Epel 2009). Норма реакции представляет собой сводные показатели диапазона фенотипов, изменяющихся количественно или качественно, и проявляемых организмами данного генотипа при различных условиях окружающей среды. Когда в норме реакции проявляются прерывистая вариация или двухвалентные фенотипы (а не количественные, непрерывные вариации), её часто называют полифенизмом (рис. 6).

Рисунок 6: Полифенизм цвета у американских гусениц пяденицы березовой (Biston betularia) является примером фенотипической пластичности.
Фенотипическая пластичность неоднократно вызывала интерес у исследователей-биологов и порождала противоречия в эволюционной теории. Всестороннее изучение фенотипической пластичности проводилось в тесной связи с количественными генетическими методами и анализом фенотипического отбора, где была продемонстрирована степень пластичности в естественных популяциях и были определены оперативные меры для ее обнаружения (Scheiner 1993; Pigliucci 2001). Другие аспекты пластичности требуют различных исследовательских методик, которые позволили бы установить источники пластичности в онтогенезе, способствующие пластичности молекулярно-генетические механизмы, а также виды функций отображения, существующие между генотипом и фенотипом (Pigliucci 2001; Kirschner and Gerhart 2005: ch. 5). Последние из упомянутых аспектов – возникновение фенотипической вариации во время и после онтогенеза – находятся на пересечении развития и эволюции: как молекулярно-генетические механизмы производят (или снижают пластичность? Какие функции отображения генотипа-фенотипа распространены, какие редки? Вносит ли пластичность свой вклад в возникновение новизны в эволюции (West-Eberhard 2003)?
Чтобы экспериментально оценить эти вопросы, нам необходимо изменить ход развития с помощью манипуляции с переменными окружающей среды и пронаблюдать, как можно сформировать новый фенотип в рамках существующей пластичности организма (Kirschner and Gerhart 2005: ch. 5). Подобное манипулирование могло бы позволить определить закономерности в изменениях при помощи надежного воспроизведения конкретных экспериментальных изменений в разных режимах окружающей среды. Однако без измерения вариаций в разных режимах окружающей среды вы не сможете наблюдать фенотипическую пластичность. Эти измерения необходимы, чтобы документировать степень пластичности и ее закономерности для конкретного признака, как, например, качественно различных морф. Чтобы оценить значимость фенотипической пластичности для эволюции, требуются ответить на вопросы о том, где возникает пластичность, как молекулярно-генетические механизмы участвуют в пластичности и какие достигаются генотип-фенотипические соотношения.
Использование стадий развития намеренно игнорирует вариации, связанные с фенотипической пластичностью. Животные и растения выращиваются в условиях стабильной окружающей среды, что и позволяет воспроизводить стадии в разных лабораторных условиях. А вариации часто рассматриваются как шум, который, если нам требуется понять работу развития, нужно уменьшить или устранить (Frankino and Raff 2004). Эта практика также поощряет выбор модельных организмов, проявляющих меньшую пластичность (Bolker, 1995). Лабораторное одомашнивание модельного организма может привести к уменьшению количества или типов наблюдаемых фенотипических изменений (Gu et al., 2005), хотя лабораторное одомашнивание может также и увеличивать вариации (например, путем инбридинга). Несмотря на попытки через контроль факторов окружающей среды уменьшить вариации, некоторые из них всегда остаются; они демонстрируют, что абсолютная хронология не является надежным способом измерения времени в онтогенезе, равно как не являются надежными ни момент начала, ни завершения различных частей онтогенеза (Mabee et al., 2000; Sheil and Greenbaum 2005). Стадии развития позволяют эффективно игнорировать эту неподатливую вариативность при помощи суждений об эмбриональной типичности. Нормальные стадии также включают в себя предположения о каузальных связях между различными процессами в последовательностях стадий (Minelli 2003: ch. 4). Как только конструирование этих стадий завершено, их можно использовать в качестве визуального стандарта, который позволяет распознавать и описывать вариацию как отклонение от нормы (DiTeresi 2010). Однако, что более типично, игнорируемые при построении этих стадий вариации также игнорируются и в рутинных обсуждениях стадий в повседневных исследовательских контекстах (Frankino and Raff 2004).
Исследование развития с помощью выделения нормальных стадий выполняют ряд целей, связанных с описательными и объяснительными проектами, в которых биологи развития принимают непосредственное участие (Kimmel et al., 1995). Исследование развития через его стадии возможность измерять экспериментальную репликацию, обеспечивают последовательную и прозрачную коммуникацию между исследователями, – особенно если стадии основаны на широко наблюдаемых морфологических признаках, – содействуют более точному прогнозированию феноменов развития, а также помогают сравнивать виды и обобщать их. В качестве идеализации онтогенеза нормальные стадии позволяют создать классификацию событий развития, имеющую всесторонний характер: надлежащим образом сортирует их по величине и достаточно однородным стадиям, устанавливает достаточно четкие границы между стадиями и обеспечивает стабильность в разных исследовательских условиях (Dupré 2001). В совокупности все это создает условия для более точных объяснений в рамках подходов отдельных дисциплин (Griesemer 1996). Идеализации также могут облегчить процедуры абстрагирования и обобщения: и то, и другое является частью экстраполяции результатов из исследовательского контекста конкретного модельного организма в другие области (Steel 2008; см. Раздел 4 и 5.1).
Исследование развития с помощью выделения нормальных стадий имеют различные недостатки, сопровождающие достижение упомянутых исследовательских и объяснительных целей. Ключевые морфологические индикаторы иногда перекрывают стадии; терминология, полезная для одной задачи, может вводить в заблуждение при использовании для другой; конкретные термины могут вводить в заблуждение при межвидовом сравнении; требуемая для продолжения наблюдений манипуляция над эмбрионом может иметь каузальное воздействие на онтогенез. Избегание изменчивости в показателях стадии может привести к игнорированию значимости конкретной вариации или, по крайней мере, служить основанием для ее минимизации.
Таким образом, существуют веские основания принять нормальные стадии для периодизации онтогенеза модельного организма, и они помогают объяснить, почему продолжающееся использование стадий приводит к эмпирическому успеху. В то же время бывают случаи, когда деление на стадии включает идеализацию – решения о намеренно неверной репрезентации конкретных видов вариаций. Подобно другим стандартным (успешным) практикам в науке, метод выделения нормальных стадий часто воспринимаются как нечто само собой разумеющееся, а это означает, что следствия из их систематических ошибок игнорируются (Wimsatt 1980), некоторые из которых имеют отношение к эволюционным вопросам (например, систематически недооценивается степень вариативности в популяции). Это важно признать, поскольку успех периодизации не зависит от потенциальной способности «смягчать» наши идеализации; периодизации нельзя постепенно скорректировать, чтобы сделать их менее идеализированными. Вместо этого конструируются новые периодизации, использующиеся вместе с уже существующими, поскольку разные идеализации включают в себя разные суждения о типичности, которые, в свою очередь, служат разнообразным описательным и объяснительным целям. В дополнение к систематическим ошибкам, связанным с выделением стадий развития, большинство модельных организмов мало что говорят нам о том, как эффекты окружающей среды модулируют или сочетаются с генетическими или другими факторами в развитии, что в итоге мешает поиску информации о механизмах, лежащих в основе норм реакции. Короткое время жизни поколения (generation time) и быстрое развитие тесно взаимосвязаны с нечувствительностью к условиям окружающей среды с помощью различных механизмов вроде формирования пред-паттернов (Bolker, 1995).
Напряжение между конкретной практикой выделения стадий развития в модельных организмах и выявлением значимости вариаций (происходящих в силу фенотипической пластичности) для эволюции можно переформулировать в виде аргумента:
Вариация, обусловленная фенотипической пластичностью, является нормальной особенностью онтогенеза.
Выделение стадий развития модельных организмов намеренно занижает вариативность онтогенеза, связанную с воздействием переменных окружающей среды (например, фенотипической пластичности), строго ограничивая диапазон значений для переменных окружающей среды, а также устраняя вариации в признаках, используемых для установления всеобъемлющей периодизации.
Следовательно, использование модельных организмов со специфицированным стадиями развития затруднит, если не сделает невозможным, наблюдение за изменениями, обусловленными фенотипической пластичностью.
Хотя это напряжение присутствует даже в том случае, когда основное внимание уделяется не вопросам эволюции (иногда поощряя биологов развития считать отсутствие доказательств доказательством незначительности фенотипической пластичности для развития), оно лишь усугубляется для исследователей эволюции. Протоколирование паттернов вариаций – вот что требуется для измерения эволюционного значения фенотипической пластичности. Практика выделения стадий развития у модельных организмов может помешать нам вынести как положительную, так и отрицательную оценку. Выделение стадий развития в сочетании со свойствами модельных организмов, как правило, способствует негативной оценке эволюционной значимости фенотипической пластичности, поскольку вариация не проявляется и не документируется, а, следовательно, вряд ли будет считаться существенной. Идеализации, включающие нормальные стадии, препятствуют тщательному экспериментальному исследованию фенотипической пластичности, что, в свою очередь, препятствует определению ее эволюционной значимости.
Последствия этой напряженности для стыковки областей биологии развития и эволюции двояки. Во-первых, самые мощные экспериментальные системы для изучения развития настроены так, чтобы минимизировать вариации, которые могут иметь решающее значение для понимания того, как эволюционные процессы происходят в природе. Во-вторых, если эволюционные исследования концентрируются вокруг признака, признанного типическим для выделения временных сегментов, которые мы называем стадиями, то бо́льшая часть вариаций этого признака концептуально отбрасывается, чтобы сделать модельный организм поддающимся экспериментированию. [ 21 ]
Выявление недостатков, сопровождающих используемые для изучения развития стратегии идеализации, не должно парализовать наши усилия. Это побуждает нас продумать то, как мы можем справиться с этими вызовами (Love 2006). Мы можем выстроить принципиальную перспективу того, как справиться с этими вызовами, добавив 3 следующие предпосылки:
Теоретические стратегии, использующиеся в идеализации, такие как (2), необходимы для успешного проведения биологических исследований онтогенеза.
Следовательно, компенсационная тактика должна быть выбрана так, чтобы точечно устранять возникающие в результате использования идеализаций слепые пятна.
Учитывая (1) – (3), компенсационная тактика должна иметь дело со следствиями игнорирования обусловленной фенотипической пластичностью вариации, которая возникает в результате выделения стадий развития у модельных организмов.
По меньшей мере две компенсационные тактики могут продвинуть наблюдения за обусловленной фенотипической пластичностью вариацией, которая обычно игнорируется при построении стадий развития для модельных организмов: тактики использования разнообразных модельных организмов и принятия альтернативных периодизаций.
Вариация будет часто наблюдаться у нестандартных модельных организмов: у экспериментальных организмов, вокруг которых не выстроено больших сообществ, с меньшей вероятностью были выделены формальные стадии эмбрионального развития, поэтому эффекты идеализации не будут воздействовать на фенотипическую пластичность. В свою очередь, исследователи стали более чувствительны к замалчиванию этих идеализаций при изучении стандартных моделей. В дальнейшем стадии могут использоваться в качестве визуальных стандартов для определения вариации как отклонения от нормы и, тем самым, характеризовать закономерности изменчивости. [ 22 ]
Вторая компенсационная тактика – принятие альтернативных периодизаций. Она включает в себя выбор разных признаков для создания новых временных членений, тем самым облегчая наблюдение за вариацией характеристик, ранее стабилизированных при периодизации нормальных стадий. Эти альтернативные периодизации часто делят подмножество событий развития в соответствии с процессами или ориентирами, отличающимися от тех, которые используются для построения нормальных стадий. Они могут не связываться один-к-одному с существующими нормальными стадиями, особенно если будут охватывать события за пределами траектории от оплодотворения до половозрелой взрослой особи. Подобное отсутствие изоморфизма между периодами также будет проявляться при использовании различных способов изменения времени, будь то последовательность (упорядочение событий) или продолжительность (последовательность определенных интервалов); кроме того, придется решать, как будут измеряться последовательности или длительности – относительно друг друга, или по внешнему стандарту, такому как абсолютная хронология (Reiss 2003, Colbert and Rowe 2008). Эти несовместимости не позволят ассимилировать альтернативные периодизации в единую всеобъемлющую схему стадий. Идеализация имеет место во всех этих случаях, поэтому каждая новая периодизация вынуждена игнорировать некоторые виды вариаций. Но альтернативные периодизации требуют выбрать другие признаки для стабилизации и типизации при определении своих временных членений, а это означает, что мы сможем наблюдать иные виды вариаций, нежели наблюдали ранее. [ 23 ]
Компенсационная тактика использования разнообразных модельных организмов и принятия альтернативных периодизаций может концептуально подходить для устранения сложностей с влиянием практики выделения стадий развития на обнаружение фенотипической пластичности; впрочем, это не устраняет все связанные с ней затраты (человеческие, финансовые и др.) или споры. Преимущества единой, всесторонней периодизации для общего модельного организма (например, нормальные стадии рыбок данио) следует оценивать с учетом преимуществ альтернативных, процесс-специализированных периодизаций. Однако, открыто и критически изучая эти практики в отношении интересующего явления, признавая преимущества и недостатки использования идеализаций, биологи развития и эволюционные биологи имеют больше возможностей для систематического описания и подробного объяснения биологических явлений.
Заключение
В этой статье мы представили лишь небольшую часть важных и перспективных сюжетов для рефлексии над эпистемологией биологии развития. О каждом из вышеупомянутых разделов биологии развития можно было бы сказать гораздо больше – например, подробнее проанализировать функционирование нормальных стадий в качестве типов (DiTeresi 2010). Кроме того, было мало сказано об использовании доказательств в экспериментах в области биологии развития, или различиях между подтверждающими и исследовательскими экспериментами (Hall 2005, Waters 2007b); также я не рассматривал роль метафор и моделей, характеризующих ключевые практики в биологии развития (Fagan 2013; Keller, 2002). Последние были тщательно проанализированы из-за повышенного внимания к деталям конкретных исследовательских программ. Наконец, ничего не было сказано о метафизических выводах по поводу явлений развития (ключевых для метафизики Аристотеля). Понятия потенциальности крайне естественны в биологии развития (например, плюрипотентность стволовых клеток или потенциал зародышевого слоя для получения различных видов тканевых линий). Это касается того, как мы понимаем предрасположенности (см. статью dispositions), поскольку условия запуска (triggering conditions) часто сложны и многократно реализованы (включая проявления без триггера); а также того факта, что клетки демонстрируют предрасположенности с несколькими возможными проявлениями (типами клеток) в специфических последовательных упорядочиваниях (см. предстоящую публикацию Hüttemann и Kaiser). Таким образом, биология развития демонстрирует не только богатый массив материальных и концептуальных практик, которые могут быть проанализированы для лучшего понимания способов научного теоретизирования, фигурирующих в экспериментальной науке о жизни, но и указывает на новые идеи для метафизики, особенно когда это начинание явным образом учитывает вклад эмпирически успешных наук.
Примечания к Биологии развития
1. Мы не рассматриваем процесс развития у растений из-за ограниченного объема статьи. Впрочем, это обстоятельство также отражает давнее предубеждение о том, как следует представлять ученикам и студентам феномены биологии развития.
2. Некоторые многоклеточные животные не имеют всех трех зародышевых слоев ткани. Например, у книдарий [стрекающих кишечнополостных], таких как медузы и кораллы, мезодермы нет.
3. Эмпирический анализ использования слова «теория» в научном дискурсе демонстрирует его колоссальную гетерогенность. В исследовании 781 статей, опубликованных в журнале Science в течение года (Overton 2013), слово «теория» не встречается среди ключевых слов 27 статей, относящихся к биологии развития, тогда как представлено более чем в 40% из 34 статей, относящихся к эволюции.
4. Минелли утверждал, что нам нужно «всеобъемлющее теоретическое описание развития» с принципом «инерции развития», аналогичным принципу инерции в механике Ньютона (Minelli 2011a, b). Мотив здесь состоит в том, чтобы поставить биологию развития на ту же теоретическую основу, на которой держится эволюционная биология.
5. «Регенерация обязательно включает в себя региональную спецификацию паттерна и, поскольку соответствующие взаимодействия являются локальными, это означает, что зашифрованная информация о паттерне должна присутствовать во всей зрелой структуре. Это иногда называют позиционной информацией. Данный термин некогда широко использовался для указания на участвующие в развитии паттерны информации, но он стал выходить из употребления по мере прояснения его молекулярной основы» (Slack 2013: 385, курсив автора).
6. Некоторые из этих переменных также помогают структурировать проблемы в других научных дисциплинах (например, абстракция и связность), но конкретно эта комбинация переменных структурирует биологию развития – особенно сопоставление временной структуры и пространственного состава.
7. Следует подчеркнуть, что подавляющее большинство исследователей, работающих в области биологии развития, предпочитают генетические объяснения и связанные с ними методологии. Они подходят настолько хорошо, что некоторые биологи развития даже не осведомлены о физических объяснениях или связанных с ними методологиях.
8. Теория систем развития (Developmental Systems Theory, DST) – это область, где попытки опровержения этих утверждений были наиболее заметны (обзор соответствующих соображений см. в: Griffiths and Stotz 2013). Важно отметить, что генетические объяснения, как уже было обозначено, совместимы с каузальными факторами, оказывающими влияние во время эмбриогенеза, весьма разнообразны и демонстрируют распределенную каузальную специфичность. В данной статье мы не пытаемся вынести решение о споре между Уотерсом (Waters 2007a) и сторонниками DST относительно того, имеют ли последовательности ДНК наибольшее влияние в биологических процессах.
9. «Молекулярная метаморфоза нашего понимания эмбриологии опирается на идентификацию генов, контролирующих развитие» (Fraser and Harland 2000: 47).
10. «Геном содержит программу инструкций по созданию организма, – порождающей программы – которая определяет, где и когда синтезируются различные белки, и, таким образом, контролирует поведение клетки» (Wolpert et al., 2010: 29).
11. Важно отметить, что критические замечания часто встречаются в том же контексте, где были выдвинуты серьезные утверждения о различительной генетической активности. Например:
Поскольку все ключевые этапы развития отражают изменения в активности генов, некто захотел бы говорить о развитии просто с точки зрения механизмов контроля экспрессии генов. Но это было бы крайне ошибочно. … Говорить только в терминах генов означает игнорировать важнейшие аспекты клеточной биологии. (Wolpert et al., 1998: 15)
12. Было бы ... преувеличением видеть в каждой кости не более, чем результат непосредственных и прямых физических или механических условий; поскольку поступать подобным образом означало бы отрицать существование, в этой связи, принципа наследственности (Thompson 1992, 1942): 1022-3).
13. Понимание морфогенеза, происхождение формы (shape) анатомических структур, органов и организмов всегда было главной целью биологии развития. ... начиная с современной революции в молекулярной биологии, ее исследовательское поле было сфокусировано на общей схеме, построенной вокруг регуляции генов, сигнальных молекул и факторов транскрипции. … Однако в последнее время исследователи снова стали признавать, что для понимания морфогенеза в трех измерениях необходимо объединить молекулярные представления (гены и морфогены) со знанием физических процессов (переноса, деформации и течения), порожденных растущими ткани (Savin et al., 2011: 57).
14. Кроме того, объединение объяснений, сочетающее генетические и физические факторы влияния, активно изучается в применении к эволюционным феноменам – таким, как происхождение новизны в эволюции в течение Кембрийского взрыва (Love and Lugar 2013).
15. «Мотив для их изучения заключается не просто в желании понять, как развивается это конкретное животное, а в том, чтобы использовать его в качестве примера развития всех животных» (Slack 2006: 61).
16. Другой ответ на критику репрезентативности, не важно, касается ли она феноменов или механизмов развития, опирается на прагматические факторы: доступность и стоимость; легкость достижения всех стадий развития и микроманипуляции; а также генетика и генетические карты (Slack 2006: ch. 6). Доступные для экспериментального анализа манипуляционные техники, большинство из которых являются молекулярными, снижают, если не устраняют, беспокойство по поводу нерепрезентативности (Davis 2004, Rubin 1988). Экспериментальный инструментарий молекулярной биологии (например, клонирование генов, гибридизация in situ, интерференция РНК и др.), фокусирующийся на молекулярных факторах влияния (раздел 3.1), лучше всего подходит для модельных организмов, и полученные посредством манипуляций каузальные знания могут в некотором смысле компенсировать нерепрезентативность (см. дополнение: «Модельные организмы и манипуляция»).
17. «Модельные организмы служат моделями для целых, неповрежденных организмов ... для целого ряда систем и процессов, происходящих в живых организмах, включая генетику, развитие, физиологию, эволюцию и экологию» (Ankeny and Leonelli 2011: 318).
18. Несколько дополнительных примеров:
Структурная и функциональная гомология между утиным и куриным гамма-интерфероном (Huang et al., 2001);
Фазическая холинергическая сигнализация в гиппокампе: функциональная гомология с неокортексом? (Gulledge и Kawaguchi 2007);
Функциональная гомология между дрожжами piD261 / Bud32 и человеческим PRPK: и фосфорилированный p53, и PRPK частично восполняют дефицит piD261 / Bud32 (Facchin et al., 2003).
19. Мы можем усилить это принятие активность-функции, изучив забытое пред-дарвиновское различие в отношении биологических функций: использование vs активность (Furley and Wilkie 1984: 58-69). Функциональное использование физиологического признака репрезентирует то, для чего он есть, тогда как функциональная активность физиологического признака отображает то, что он делает. В основе конкретного использования может лежать множество видов активности, и одна активность может обслуживать несколько видов использования; термины не имеют эквивалентных расширений. Они также не эквивалентны эпистемологически, поскольку мы можем знать, что нечто делает, но не то, для чего оно предназначено; или, иначе, мы можем знать, для чего предназначено нечто, но не то, как оно этого достигает. Каузальная роль, жизнеспособность и селекционный эффект – все это функции-использования .
20. Биохимическая функция этих генов в высокой степени сохраняется в течение эволюционного времени, в то время как их функция в развитии изменяется относительно свободно. Это неотъемлемое свойство регуляторных генов развития позволяет им независимо кооптироваться, чтобы функционировать в структурах, явно имеющих независимое эволюционное происхождение (Abouheif 1997: 407).
21. Например, стандартная периодизация пост-эмбрионального онтогенеза у членистоногих является препятствием для эволюционного анализа эволюции времени линьки, поскольку обычная периодизация конструируется через интервалы от одной линьки до другой после вылупления, которые затем группируются в стадии (личинка, куколка и имаго для насекомых), независимо от различий в сроках линьки (Minelli et al., 2006).
22. Хорошим примером здесь является исследование морфологии рога жука-навозника, которое предоставило некоторые из наиболее убедительных доказательств эволюционного значения фенотипической пластичности (Emlen 2000; Moczek and Nagy 2005, Moczek 2008). Концентрация на внутри- и межвидовой вариации специфических морфологических признаков (рогов) у насекомых, относящихся к тому же роду, что и навозный жук, означает, что нормальные стадии для одного конкретного вида не были построены с использованием времени прорезания или других характеристик онтогенеза рога. Как следствие, изменчивость этих признаков не была сведена к минимуму и остается актуальной в контексте экспериментальных наблюдений за развитием.
23. Например, чтобы более точно изучить эволюцию развития крыла, Рид и его коллеги (Reed et al., 2007) выделили стадии онтогенеза для конечного возраста диска крыла у обыкновенной бабочки Junonia coenia, поскольку время событий, происходящих с личинкой, относительно отделено от времени развития диска крыла, и, следовательно, временные измерения личиночного онтогенеза не имеют тесной корреляции с состоянием развития крыловых дисков. Это прямым образом поспособствовало изучению феномена фенотипической пластичности:
«Некоторые таинственные изменения, впрочем, могут проявляться как в плане развития, так и в физическом плане, но они просто-напросто не влияют на явные или доступные исследователям фенотипы». (Reed et al., 2007: 2)
ДОПОЛНИТЕЛЬНЫЙ МАТЕРИАЛ К РАЗДЕЛУ 4:
Модельные организмы и манипуляция
Поскольку в биологии развития модельные организмы используются для экспериментального вмешательства, вопросы о репрезентативности (является ли механизм формирования сомитов у рыбок данио хорошим эрзац-образцом механизма образования сомитов у позвоночных?) необходимо сопоставлять с вопросами манипуляции (насколько легко и сколь разнообразно можно манипулировать формированием сомитов у рыбок данио в эксперименте?). Когда Слэк описывает преимущества стандартных моделей развития животных, манипуляция выступает приоритетом номер один:
«Каждый из шести модельных видов был выбран потому, что у них есть некоторые специфические экспериментальные преимущества для исследований в области биологии развития». (Slack 2006: 61, курсив автора)
Может показаться, что критерий манипулятивности не зависит от критерия репрезентативности, однако между ними существуют потенциальные пересечения. Например, если модельный организм выступает представителем сразу множества механизмов развития, но не всеми из них легко манипулировать, это может повлиять на выбор и использование данного организма. Поскольку многие генетические методы оказались неприменимы к куриным эмбрионам, роль последних в качестве модельных организмов в течение последнего десятилетия снизилась, хотя они все еще широко используются, что оправдано с точки зрения манипуляций. Манипулятивность в некоторых обстоятельствах может даже быть важнее репрезентативности.
Одно из важных различий пролегает между тем, насколько легко инициировать экспериментальный режим с данным модельным организмом («первоначальная настройка»), и насколько легко к ним применять различные экспериментальные методы («исполнение»). Для того, чтобы сделать некий организм модельным, требуется много ресурсов. Некоторые из этих организмов имеют долгую историю (цыпленок), тогда как другие стали модельными организмами совсем недавно (данио). Но во всех случаях для того, чтобы это осуществить, необходимо вложить значительные инвестиции времени, энергии и навыков. Так, мы можем подразделить аспект первоначальной настройки на категории «доступности и стоимости» и другой категории, относящейся к «инфраструктурным предварительным условиям». Если модельный организм размножается нечасто и нерегулярно, а периоды его беременности очень велики, ученым будет сложно собрать необходимый запас эмбрионов на разных стадиях исследования развития. Даже при соблюдении данных условий, это окажется бесполезным, если сопутствующие траты невозможно оформить в стандартизованной шкале расходов (которая могла бы быть предусмотрена в бюджете исследования в обычном режиме). Доступность и стоимость явным образом взаимосвязаны с инфраструктурой; возможность иметь эмбрионы животных под рукой при низкой удельной стоимости зависит от имеющейся инфраструктуры (например, резервуаров для рыбок данио). Требуемая инфраструктура строится по частям, по мере того как организм выбирается и используется; часто это служит способом уменьшить затраты или обеспечить одинаковую доступность (например, садки для косяков рыб). Репрезентативная привлекательность новых модельных организмов и способность адаптировать существующие экспериментальные протоколы не заменяет соображений доступности/стоимости и инфраструктурных издержек (Crotty and Gann 2009, Slack 2009).
Второй аспект критерия манипуляции – это исполнение; его можно подразделить на «ассортимент» (каково возможное количество различных экспериментальных манипуляций?) и «скорость» (сколь быстро можно выполнить эти экспериментальные манипуляции, чтобы получить данные?). Эти подразделения дают понять, что исполнение зависит от соображений, сформулированных на этапе первоначальной настройки. У исследователя не будет возможности освоить весь ассортимент экспериментальных манипуляций, если организмы недоступны и их стоимость слишком высока или же если отсутствует соответствующая инфраструктура: например, если мы не можем разместить достаточное количество животных, нужное для таргетного разведения (targeted breeding), или если мы не можем инвестировать в секвенирование генома. Но ассортимент и скорость связаны с другими практическими предварительными условиями (например, легкостью получения организма на всех этапах развития и микроманипуляции). Без полного доступа определенные виды экспериментальных вмешательств будут просто невозможны (т.е. ассортимент уменьшается). Если микроманипуляция того или иного рода сложна, скорость исполнения уменьшится. Ассортимент и скорость связаны друг с другом, потому что один метод вмешательства может быть выполнен легко и в разумные сроки, тогда как другой – нет. В целом, модельные организмы выбираются и используются таким образом, чтобы максимизировать как ассортимент, так и скорость, однако существует много компромиссов. Микрохирургические эмбриональные манипуляции с цыплятами можно считать быстрыми по сравнению с манипуляциями с Drosophila , но особями Drosophila гораздо легче (и быстрее) манипулировать генетически.
Хотя оценки различных аспектов критерия репрезентации отличаются от оценок аспектов критерия манипуляции, эти два критерия могут, в сущности, обуславливать друг друга. Репрезентационная способность организма-модели может быть не связана с его податливостью манипуляции (и наоборот), безотносительно того, идет ли речь о явлениях или механизмах. Впрочем, взаимообусловленность может быть высока в том случае, когда высокая подверженность манипуляции усиливает или уменьшает репрезентативность. В большинстве обсуждений подчеркивалось, что лабораторное одомашнивании снижает изменчивости, когда очищенные генетические линии либо стандартизированные экологические режимы снижают количество естественных изменений организма или меняют другие его нормальные функции (Bolker, 1995, Frankino and Raff 2004; Gu et al., 2005; Ankeny 2009). Хотя полностью признано, что подобные типы эффектов присутствуют, во многом игнорируется противоположная возможность – то, насколько манипуляции могут улучшить репрезентативность или создать более удачные модели явлений и механизмов развития. Интересным следствием того пристального внимания, которое уделяется различным аспектам манипуляции и репрезентации, является возможность привлечения новых модельных организмов. Считается, например, что микробные виды менее репрезентативны в качестве феноменов развития – но они могут быть спроектированы таким образом, чтобы экспонировать эти феномены, т.к. микробные виды лучше удовлетворяют критериям манипуляции (Love and Travisano 2013). Более тонкое понимание эпистемологической подоплеки выбора и использования модельного организма в биологии развития позволит предложить новые возможности для эмпирических исследований, которые дополнят существующие способы практического применения модельного организма в биологии развития.
Библиография
Abouheif, E., 1997, “Developmental genetics and homology: a hierarchical approach”, Trends in Ecology and Evolution, 12: 405–408.
Abouheif, E., M. Akam, W.J. Dickinson, P.W.H. Holland, A. Meyer, N.H. Patel, R.A. Raff, V.L. Roth, and G.A. Wray, 1997, “Homology and developmental genes”, Trends in Genetics, 13: 432–433.
Ankeny, R.A., 2009, “Model organisms as fictions”, in Fictions in Science: Philosophical Essays on Modeling and Idealization, M. Suárez (ed.), 193–204. New York and London: Routledge, Tayor & Francis Group.
–––, 2012, “Detecting themes and variations: the use of cases in developmental biology”, Philosophy of Science, 79: 644–654.
Ankeny, R.A., and S. Leonelli, 2011, “What’s so special about model organisms?”, Studies in History and Philosophy of Science, 42: 313–323.
Bechtel, W, and R. Richardson, 1993, Discovering Complexity: Decomposition and Localization as Strategies in Scientific Research, Princeton: Princeton University Press.
Berrill, N.J., 1961, Growth, Development, and Pattern, San Francisco: W.H. Freeman and Company.
Bier, E., and W. McGinnis, 2003, “Model organisms in the study of development and disease”, in Molecular Basis of Inborn Errors of Development, C.J. Epstein, R.P. Erickson, and A. Wynshaw-Boris (eds.), 25–45. New York: Oxford University Press.
Bolker, J.A., 1995, “Model systems in developmental biology”, BioEssays, 17: 451–455.
–––, 2009, “Exemplary and surrogate models: Two modes of representation in biology”, Perspectives in Biology and Medicine, 52: 485–499.
Brand, T., 2003, “Heart development: molecular insights into cardiac specification”, Developmental Biology, 258: 1–19.
Brigandt, I., 2013, “Systems biology and the integration of mechanistic explanation and mathematical explanation”, Studies in History and Philosophy of Biological and Biomedical Sciences, 44: 477–492.
Brigandt, I., and A.C. Love, 2012, “Conceptualizing evolutionary novelty: Moving beyond definitional debates”, Journal of Experimental Zoology (Mol Dev Evol), 318B: 417–427.
Brouzés, E., and E. Farge, 2004, “Interplay of mechanical deformation and patterned gene expression in developing embryos”, Current Opinion in Genetics & Development, 14: 367–374.
Colbert, M.W., and T. Rowe, 2008, “Ontogenetic Sequence Analysis: using parsimony to characterize developmental sequences and sequence polymorphism”, Journal of Experimental Zoology (Mol Dev Evol), 310B: 398–416.
Craver, C.F., 2007, Explaining the Brain: Mechanisms and the Mosaic Unity of Neuroscience, New York: Oxford University Press.
Crotty, D.A., and A. Gann, 2009, Emerging Model Organisms: A Laboratory Manual, Volume 1, Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Darden, L., 2006, Reasoning in Biological Discoveries: Essays on Mechanisms, Interfield Relations, and Anomaly Resolution, New York: Cambridge University Press.
Davidson, E.H., 2006, The Regulatory Genome: Gene Regulatory Networks in Development and Evolution, San Diego: Academic Press.
Davies, J.A., 2005, Mechanisms of Morphogenesis: The Creation of Biological Form, San Diego, CA: Elsevier Academic Press.
Davis, R.H., 2004, “The age of model organisms”, Nature Reviews Genetics, 5: 69–76.
DeWitt, T.J., and S.M. Scheiner, 2004, Phenotypic Plasticity: Functional and Conceptual Approaches, New York: Oxford University Press.
DiTeresi, C.A., 2010, “Taming Variation: Typological Thinking and Scientific Practice in Developmental Biology”, University of Chicago, Chicago. PhD Thesis.
Dupré, J., 2001, “In defence of classification”, Studies the History and Philosophy of Biological and Biomedical Sciences, 32: 203–219.
Ellis, R.J., 2001, “Macromolecular crowding: obvious but underappreciated”, Trends in Biochemical Sciences, 26: 597–604.
Emlen, D.J., 2000, “Integrating development with evolution: A case study with beetle horns”, BioScience, 50: 403–418.
Facchin, S., R. Lopreiato, M. Ruzzene, O. Marin, G. Sartori, C. Götz, M. Montenarh, G. Carignani, and L.A. Pinna, 2003, “Functional homology between yeast piD261/Bud32 and human PRPK: both phosphorylate p53 and PRPK partially complements piD261/Bud32 deficiency”, FEBS Letters, 549: 63–66.
Fagan, M.B., 2013, Philosophy of Stem Cell Biology: Knowledge in Flesh and Blood, London: Palgrave Macmillan.
Forgacs, G., and S.A. Newman, 2005, Biological Physics of the Developing Embryo, New York: Cambridge University Press.
Frankino, W.A., and R.A. Raff, 2004, “Evolutionary importance and pattern of phenotypic plasticity”, in DeWitt and Scheiner 2004: 64–81.
Fraser, S.E., and R.M. Harland, 2000, “The molecular metamorphosis of experimental embryology”, Cell, 100: 41–55.
Furley, D., and J.S. Wilkie, 1984, Galen: On Respiration and the Arteries, Princeton: Princeton University Press.
Gajewski, K., and R.A. Schulz, 2002, “Comparative genetics of heart development: conserved cardiogenic factors in Drosophila and vertebrates”, in Cardiac Development, B. Ostadal, M. Nagano, and N.S. Dhalla (eds.), 1–23. Boston: Kluwer Academic Publishers.
Gerhart, J., and M. Kirschner, 2007, “The theory of facilitated variation”, Proceedings of the National Academy of Sciences USA, 104: 8582–8589.
Ghiselin, M.T., 2005, “Homology as a relation of correspondence between parts of individuals”, Theory in Biosciences, 124: 91–103.
Giere, R.N., 1999, Science Without Laws, Chicago: University of Chicago Press.
Gilbert, S.F. (ed.), 1991, A Conceptual History of Modern Embryology (Volume 7: Developmental Biology: A Comprehensive Synthesis), New York: Plenum Press.
–––, 2000 [2003, 2006, 2010], Developmental Biology, 6th edition, Sunderland, MA: Sinauer Associates, Inc., 2000; 7th edition, 2003; 8th edition, 2006; 9th edition, 2010.
Gilbert, S.F. and D. Epel, 2009, Ecological Developmental Biology: Integrating Epigenetics, Medicine, and Evolution, Sunderland, MA: Sinauer.
Greene, E., 1989, “A diet-induced developmental polymorphism in a caterpillar”, Science, 243: 643–646.
Griesemer, J.R., 1996, “Periodization and models in historical biology”, in New Perspectives on the History of Life, M.T. Ghiselin, and G. Pinna (eds.), 19–30. San Francisco: California Academy of Sciences.
Griffiths, P. and K. Stotz, 2013, Genetics and Philosophy: An Introduction, New York: Cambridge University Press.
Gu, Z., L. David, D. Petrov, T. Jones, R.W. Davis, and L.M. Steinmetz, 2005, “Elevated evolutionary rates in the laboratory strain of Saccharomyces cerevisiae”, Proceedings of the National Academy of Sciences USA, 102: 1092–1097.
Gulledge, A.T., and Y. Kawaguchi, 2007, “Phasic cholinergic signaling in the hippocampus: functional homology with the neocortex? Hippocampus, 17: 327–332”,
Hall, B.K. (ed.), 1994, Homology: The Hierarchical Basis of Comparative Biology, San Diego: Academic Press.
Hall, L.R., 2005, “Exploratory experiments”, Philosophy of Science, 72: 888–899.
Hamburger, V., 1988, The Heritage of Experimental Embryology: Hans Spemann and the Organizer, New York: Oxford University Press.
Hamburger, V., and H.L. Hamilton, 1951, “A series of normal stages in the development of the chick embryo”, Journal of Morphology, 88: 49–92.
Harvey, R.P., 2002, “Patterning the vertebrate heart”, Nature Reviews Genetics, 3: 544–556.
Hoffman, B.D., C. Grashoff, and M.A. Schwartz, 2011, “Dynamic molecular processes mediate cellular mechanotransduction”, Nature, 475: 316–323.
Hopwood, N., 2005, “Visual standards and disciplinary change: normal plates, tables and stages in embryology”, History of Science, 43: 239–303.
–––, 2007, “A history of normal plates, tables and stages in vertebrate embryology”, International Journal of Developmental Biology, 51: 1–26.
Horder, T.J., J.A. Witkowski, and C.C. Wylie (eds), 1986, A History of Embryology, Cambridge: Cambridge University Press.
Hove, J.R., R.W. Köster, A.S. Forouhar, G. Acevedo-Bolton, S.E. Fraser, and M. Gharib, 2003, “Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis”, Nature, 421: 172–177.
Huang, A., C.A. Scougall, J.W. Lowenthal, A.R. Jilbert, and I. Kotlarski, 2001, “Structural and functional homology between duck and chicken interferon-gamma”, Developmental and Comparative Immunology, 25: 55–68.
Hüttemann, A. and M.I. Kaiser, forthcoming, “Potentiality in biology”, in Handbook of Potentiality, K. Engelhard and M. Quante (eds.), Berlin: Springer.
Jones, M.R., 2005, “Idealization and abstraction: a framework”, in Idealization XII: Correcting the Model. Idealization and Abstraction in the Sciences (Poznan Studies in the Philosophy of the Sciences and the Humanities, vol. 86), M.R. Jones and N. Cartwright (eds.), 173–217. Amsterdam/New York: Rodopi.
Kaplan, J.M., 2008, “Phenotypic plasticity and reaction norms”, in Sarkar and Plutynski 2008: 205–222.
Keller, E.F., 2002, Making Sense of Life: Explaining Biological Development with Models, Metaphors, and Machines, Cambridge, MA: Harvard University Press.
Kimmel, C.B., W.W. Ballard, S.R. Kimmel, B. Ullmann, and T.F. Schilling, 1995, “Stages of embryonic development of the zebrafish”, Developmental dynamics, 203: 253–310.
Kirby, M.L., 1999, “Contribution of neural crest to heart and vessel morphology”, in Heart Development, R.P. Harvey and N. Rosenthal (eds.), 179–193. San Diego: Academic Publishers.
Kirschner, M.W., and J.C. Gerhart, 2005, The Plausibility of Life: Resolving Darwin’s Dilemma, New Haven and London: Yale University Press.
Kitcher, P., 1993, The Advancement of Science: Science Without Legend, Objectivity Without Illusions, New York: Oxford University Press.
Levin, M., T. Thorlin, K.R. Robinson, T. Nogi, and M. Mercola, 2002, “Asymmetries in H+/K+ -ATPase and cell membrane potentials comprise a very early step in left-right patterning”, Cell, 111: 77–89.
Love, A.C., 2006, “Taking development seriously: who, what, where, why, when, how?”, Biology & Philosophy, 21: 575–589.
–––, 2007, “Functional homology and homology of function: biological concepts and philosophical consequences”, Biology & Philosophy, 22: 691–708.
–––, 2008a, “Explaining evolutionary innovation and novelty: criteria of explanatory adequacy and epistemological prerequisites”, Philosophy of Science, 75: 874–886.
–––, 2008b, “Explaining the ontogeny of form: philosophical issues”, in Sarkar and Plutynski 2008: 223–247.
–––, 2010a, “Idealization in evolutionary developmental investigation: a tension between phenotypic plasticity and normal stages”, Philosophical Transactions of the Royal Society B: Biological Sciences, 365: 679–690.
–––, 2010b, “Rethinking the structure of evolutionary theory for an extended synthesis”, in Evolution—The Extended Synthesis, M. Pigliucci, and G.B. Müller (eds.), 403–441. Cambridge, MA: MIT Press.
–––, 2012, “Formal and material theories in philosophy of science: A methodological interpretation”, in EPSA Philosophy of Science: Amsterdam 2009 (The European Philosophy of Science Association Proceedings, Vol. 1), H.W. de Regt, S. Okasha, and S. Hartmann (eds.), 175–185. Berlin: Springer.
–––, 2013, “Theory is as theory does: Scientific practice and theory structure in biology”, Biological Theory, 7: 325–337.
–––, 2014, “The erotetic organization of development”, in Towards a Theory of Development, A. Minelli, and T. Pradeu (eds.), 33–55. Oxford: Oxford University Press.
–––, 2015, “Evolutionary developmental biology: philosophical issues”, in Handbook of Evolutionary Thinking in the Sciences, T. Heams, P. Huneman, L. Lecointre, and M. Silberstein (eds.), 265–283, Berlin: Springer.
–––, forthcoming, “Combining genetic and physical causation in developmental explanations”, in Causal Reasoning in Biology, C.K. Waters and J. Woodward (eds.), Minnesota Studies in Philosophy of Science, Minneapolis, MN: University of Minnesota Press.
Love, A.C. and G.L. Lugar, 2013, “Dimensions of integration in interdisciplinary explanations of the origin of evolutionary novelty”, Studies in the History and Philosophy of Biological and Biomedical Sciences, 44: 537–550.
Love, A.C., and M. Travisano, 2013, “Microbes modeling ontogeny”, Biology & Philosophy, 28: 161–188.
Lynch, V.J., 2009, “Use with caution: developmental systems divergence and potential pitfalls of animal models”, Yale Journal of Biology and Medicine, 82: 53–66.
Mabee, P.M., K.L. Olmstead, and C.C. Cubbage, 2000, “An experimental study of intrspecific variation, developmental timing, and heterochrony in fishes”, Evolution, 54: 2091–2106.
Maienschein, J., 1991, Transforming Traditions in American Biology, 1880–1915, Baltimore, MD: The Johns Hopkins University Press.
–––, 2000, “Competing epistemologies and developmental biology”, in Biology and Epistemology, R. Creath and J. Maienschein (eds.), 122–137. Cambridge: Cambridge University Press.
Maienschein, J., M. Glitz, and G.E. Allen, 2005, Centennial History of the Carnegie Institution of Washington: Volume 5, The Department of Embryology, New York: Cambridge University Press.
Manak, J.R., and M.P. Scott, 1994, “A class act: conservation of homeodomain protein functions”, Development, (Supplement): 61–71.
Metscher, B.D., and P.E. Ahlberg, 1999, “Zebrafish in context: uses of a laboratory model in comparative studies”, Developmental Biology, 210: 1–14.
Mill, J.S., 1843 [1974], A System of Logic Ratiocinative and Inductive, Being a Connected View of the Principles of Evidence and the Methods of Scientific Investigation (Books I–III), in The Collected Works of John Stuart Mill, Volume VII, John M. Robson (ed.), Toronto: University of Toronto Press, London: Routledge and Kegan Paul. [Mill 1843 [1974] available online]
Miller, C.J., and L.A. Davidson, 2013, “The interplay between cell signalling and mechanics in developmntal processes”, Nature Reviews Genetics, 14: 733–744.
Minelli, A., 2003, The Development of Animal Form: Ontogeny, Morphology, and Evolution, Cambridge: Cambridge University Press.
–––, 2011a, “Animal development, an open-ended segment of life”, Biological Theory, 6: 4–15.
–––, 2011b, “A principle of developmental inertia”, in Epigenetics: Linking Genotype and Phenotype in Development and Evolution, B. Halgrímsson and B.K. Hall (eds.), 116–133. San Francisco: University of California Press.
Minelli, A., C. Brena, G. Deflorian, D. Maruzzo, and G. Fusco, 2006, “From embryo to adult-beyond the conventional periodization of arthropod development”, Development Genes and Evolution, 216: 373–383.
Mitchell, S.D., 2002, “Integrative pluralism”, Biology & Philosophy, 17: 55–70.
Moczek, A., 2008, “On the origins of novelty in development and evolution”, BioEssays, 30: 432–447.
Moczek, A.P., and L.M. Nagy, 2005, “Diverse developmental mechanisms contribute to different levels of diversity in horned beetles”, Evolution & Development, 7: 175–185.
Morgan, T.H., 1923, “The modern theory of genetics and the problem of embryonic development”, Physiological Review, 3: 603–627.
–––, 1926, “Genetics and the physiology of development”, American Naturalist, 60: 489–515.
–––, 1934, Embryology and Genetics, New York: Columbia University Press.
Moss, L., 2002, What Genes Can’t Do, Cambridge, MA: MIT Press, A Bradford Book.
Müller, G.B., 2007, “Evo-devo: extending the evolutionary synthesis”, Nature Reviews Genetics, 8: 943–949.
Nagel, E., 1961, The Structure of Science: Problems in the Logic of Scientific Explanation, New York: Harcourt, Brace & World, Inc.
Neumann-Held, E.M., and C. Rehmann-Sutter (eds.), 2006, Genes in Development: Re-reading the Molecular Paradigm, Durham and London: Duke University Press.
Newman, S.A., and R. Bhat, 2008, “Dynamical patterning modules: physico-genetic determinants of morphological development and evolution”, Physical Biology, 5: 1–14.
Nonaka, S., H. Shiratori, Y. Saijoh, and H. Hamada, 2002, “Determination of left-right patterning of the mouse embryo by artificial nodal flow”, Nature, 418: 96–99.
Nüsslein-Volhard, C., 2006, Coming to Life: How Genes Drive Development, Carlsbad, CA: Kales Press.
Olby, R.C., 1986, “Structural and dynamical explanations in the world of neglected dimensions”, in Horder, Witkowski, and Wylie 1986: 275–308.
Olson, E.N., 2006, “Gene regulatory networks in the evolution and development of the heart”, Science, 313: 1922–1927.
Oppenheimer, J.M., 1967, Essays in the History of Embryology and Biology, Cambridge, MA: MIT Press.
Overton, J.A., 2013, “‘Explain’ in scientific discourse”, Synthese, 190: 1383–1405.
Owen, R., 1843, Lectures on the Comparative Anatomy and Physiology of the Invertebrate Animals, London: Longman, Brown, Green, and Longmans.
Pabst, D.A., 2000, “To bend a dolphin: convergence of force transmission designs in Cetaceans and Scombrid fishes”, American Zoologist, 40:146–155.
Parkkinen, V-P., 2014, “Developmental explanations”, in New Directions in the Philosophy of Science: The Philosophy of Science in a European Perspective, Vol. 5, M.C. Galavotti, D. Dieks, W.J. Gonzalez, S. Hartmann, T. Uebel, and M. Weber (eds.), Berlin: Springer.
Peter, I.S., and E.H. Davidson, 2011, “A gene regulatory network controlling the embryonic specification of endoderm”, Nature, 474: 635–639.
Pigliucci, M., 2001, Phenotypic Plasticity: Beyond Nature and Nurture, Baltimore and London: The Johns Hopkins University Press.
–––, 2002, “Touchy and bushy: phenotypic plasticity and integration in response to wind stimulation in Arabidopsis thaliana”, International Journal of Plant Sciences, 163: 399–408.
Raff, R.A., 2000, “Evo-Devo: the evolution of a new discipline”, Nature Reviews Genetics, 1: 74–79.
Ranganayakulu, G., D.A. Elliott, R.P. Harvey, and E.N. Olson, 1998, “Divergent roles for NK-2 class homeobox genes in cardiogenesis in flies and mice”, Development, 125: 3037–3048.
Raya, Á., Y. Kawakami, C. Rodríguez-Esteban, M. Ibañes, D. Rasskin-Gutman, J. Rodríguez-León, D. Büscher, J.A. Feijó, and J.C.I. Belmonte, 2004, “Notch activity acts as a sensor for extracellular calcium during vertebrate left-right determination”, Nature, 427: 121–128.
Reed, R.D., P-H. Chen, and H.F Nijhout, 2007, “Cryptic variation in butterfly eyespot development: the importance of sample size in gene expression studies”, Evolution & Development, 9: 2–9.
Reiss, J.O., 2003, “Time”, in Keywords and Concepts in Evolutionary Developmental Biology, B.K. Hall, and W.M. Olson (eds.), 359–368. Cambridge, MA: Harvard University Press.
Robert, J.S., 2004, Embryology, Epigenesis, and Evolution: Taking Development Seriously, New York: Cambridge University Press.
Roe, S.A., 1981, Matter, Life, and Generation: 18th Century Embryology and the Haller-Wolff Debate, Cambridge: Cambridge University Press.
Rosenberg, A., 2006, Darwinian Reductionism: Or, How to Stop Worrying and Love Molecular Biology, Chicago: University of Chicago Press.
Rubin, G.M., 1988, Drosophila melanogaster as an experimental organism. Science, 240: 1453–1459.
Sarkar, S. and A. Plutynski (eds.), 2008, A Companion to the Philosophy of Biology, (Blackwell Companions to Philosophy), Malden, MA: Blackwell Publishers.
Savin, T., N.A. Kurpios, A.E. Shyer, P. Florescu, H. Liang, L. Mahadevan, and C. Tabin, 2011, “On the growth and form of the gut”, Nature, 476: 57–62.
Scheiner, S.M., 1993, “Genetics and evolution of phenotypic plasticity”, Annual Review of Ecology and Systematics, 24: 35–68.
Schulze, J., and E. Schierenberg, 2011, “Evolution of embryonic development in nematodes”, EvoDevo, 2: 18.
Sheil, C.A., and E. Greenbaum, 2005, “Reconsideration of skeletal development of Chelydra serpentina (Reptilia: Testudinata: Chelydridae): evidence for intraspecific variation”, Journal of Zoology, 265: 235–267.
Slack, J.M.W., 2006, Essential Developmental Biology, 2nd ed. Malden, MA: Blackwell Publishing.
–––, 2009, “Emerging market organisms”, Science, 323: 1674–1675.
–––, 2013, Essential Developmental Biology, 3rd ed. Chichester, UK: Wiley-Blackwell.
Smith, J.E.H. (ed.), 2006, The Problem of Animal Generation in Early Modern Philosophy, New York: Cambridge University Press.
–––, 2011, Divine Machines: Leibniz and the Sciences of Life, Princeton, NJ: Princeton University Press.
Sober, E., 1988, “Apportioning causal responsibility”, Journal of Philosophy, 85: 303–318.
Sommer, R.J., 2009, “The future of evo-devo: model systems and evolutionary theory”, Nature Reviews Genetics, 10: 416–422.
Srivastava, D., 2006, “Making or breaking the heart: from lineage determination to morphogenesis”, Cell, 126: 1037–1048.
Steel, D.P., 2008, Across the Boundaries: Extrapolation in Biology and Social Science, New York: Oxford University Press.
Strevens, M., 2009, Depth: An Account of Scientific Explanation, Cambridge, MA: Harvard University Press.
Thompson, D’A.W. 1992 [1942]. On Growth and Form, Complete Revised Edition. New York: Dover Publications, Inc.
Vázquez-Novelle, M.D., V. Esteban, A. Bueno, and M.P. Sacristán., 2005, “Functional homology among human and fission yeast Cdc14 phosphatases”, Journal of Biological Chemistry, 280: 29144–29150.
Von Dassow, M., J. Strother, and L.A. Davidson, 2010, “Surprisingly simple mechanical behavior of a complex embryonic tissue”, PLoS ONE, 5:e15359.
Waters, C.K., 2007a, “Causes that make a difference”, Journal of Philosophy, 104: 551–579.
–––, 2007b, “The nature and context of exploratory experimentation”, History and Philosophy of the Life Sciences, 29: 275–284.
Weber, M., 2005, Philosophy of Experimental Biology, New York: Cambridge University Press.
Weisberg, M., 2007, “Three kinds of idealization”, Journal of Philosophy, 104: 639–659.
West-Eberhard, M.J., 2003, Developmental Plasticity and Evolution, New York: Oxford University Press.
Wimsatt, W.C., 1980, “Reductionistic research strategies and their biases in the units of selection controversy”, in Scientific Discovery: Case Studies, T. Nickles (ed.), 213–259. Dordrecht: D. Reidel Publishing Company.
Wolpert, L., R. Beddington, J. Brockes, T. Jessell, P.A. Lawrence, and E.M. Meyerowitz, 1998, Principles of Development, New York: Oxford University Press.
Wolpert, L., C. Tickle, T. Jessell, P. Lawrence, E. Meyerowitz, E. Robertson, and J. Smith, 2010, Principles of Development, 4th ed. New York and Oxford: Oxford University Press.
Woodward, J., 2003, Making Things Happen: A Theory of Causal Explanation, New York: Oxford University Press.
Wouters, A., 2003, “Four notions of biological function”, Studies in the History and Philosophy of Biological and Biomedical Sciences, 34: 633–668.
–––, 2005, “The function debate in philosophy”, Acta Biotheoretica, 53: 123–151.
Wozniak, M., and C.S. Chen, 2009, “Mechanotransduction in development: a growing role for contractility”, Nature Reviews Molecular Cell Biology, 10: 34–43.









