




Философия нейронаук
- До и после нейрофилософии
- Элиминативный материализм и нейронизированная философия
- Нейронаука и психосемантика
- Объяснённое сознание?
- Локализация когнитивных функций: от изучения поражений до функциональной нейровизуализации
- Результат коэволюционной исследовательской идеологии: фокусировка философии на когнитивной и вычислительной нейробиологии
- Достижения в философии нейронауки
- Достижения за второе десятилетие XXI века
- Bibliography
Впервые опубликовано 7 июня 1999 г .; Основное переработано 6 августа 2019 г.
За последние четыре десятилетия философия становилась все более «частной». Фокус внимания сместился от общих, присущих практике черт, проблем и затруднениям научным характерным для конкретных дисциплин.
Философия нейронаук‹1› - один из естественных результатов данного процесса. Стимулом к развитию этой также послужил значительный рост достижений в качестве нейронауках.
Когнитивная и вычислительная нейробиология продолжает рассматривать вопросы, традиционно рассматриваемые в гуманитарных науках, включая природу сознания, действия, знания и нормативности. Клеточная, молекулярная и поведенческая нейробиология, использующие экспериментальные модели на животных, все чаще исследуют сферу когнитивной нейробиологии. Эмпирические открытия, соответствующие структуры и функционирование мозга, вызывают подходы, которые позволяют «натуралистические» программы разработать в деталях, к себе выходя за пределы абстрактных философских соображений.
Уже два десятилетия в релевантной литературе присутствует различие между «философией нейронаук» и «нейрофилософией»: первая из них касается основополагающих проблем в нейронауках, последняя - применения нейробиологических понятий к традиционным философским вопросам. Изучение различных концепций репрезентации, используемых в нейронаучных теориях, является примером первого. Изучение следствий неврологических синдромов, следующих для концепции «единого Я», является примером последнего. В настоящей статье мы продолжим и приведим примеры как философии нейронаук, так и нейрофилософии. Как это имело место и в истории полевых исследований, работы в каждой из этих двух областей разбросаны по всем нижеследующим разделам.
На всём протяжении статьи мы будем пытаться указать, к какой из двух областей относится значимая работа, когда это не представляется очевидным.
Один из наиболее захватывающих условий работы в философии нейронаук или нейрофилософии - это постоянный элемент удивления. Каждая из зависимостей зависит от развития нейронаук. То, что было научной фантастикой в прошлом году, в этом году становится научной реальностью. Но особенность лишь усложняет задачу около десятилетия энциклопедической статьи: научные детали, о которых философы размышляли в прошлых обновлениях, сегодня прочтены как прискорбно устаревшие.
Тем не менее, найдутся и те читатели, которые захотят познакомиться с историей продолжающейся развиваться области. Наше решение данной дилеммы состояло в том, чтобы сохранить предыдущие дискуссии, отразив историю области, однако также добавить более свежие научные и философские обновления, - причём не только в разделах данной статьи, недавних исследований, но и в разделах, посвящённых ранним дискуссиям. Хотя такое решение не претендует на идеальность, оно позволяет сохранить что-то из истории философии нейронаук и нейрофилософии на фоне постоянного прогресса в науках, от которых зависят эти философские области.
До и после нейрофилософии
Исторически, влияние нейронаучных открытий на детали материалистических программ в философии сознания было небольшое. «Нейронаучная среда» второй половины прошлого века для философов проблематичным принятие субстанционального дуализма в отношении сознания. Однако даже типовые («тип-тип») теории тождества или теории тождества «центрального состояния» («центральное состояние»), получая широкое распространение в конце 1950-х годов (Place 1956; Smart 1959), опирались на некоторые реальные детали устанавливающих нейронаук . Обращаясь к ранним примерам, вспомним излюбленное утверждение о психоневральном тождестве: «боль идентична активации C-волокон».
Как оказалось, «С-волокна» связаны только с одним аспектом передачи боли (Hardcastle 1997). Ранние теоретики тождества не делали акцент на гипотезах психоневральной идентичности. Их «нейро-» термины, по общему признанию, были своего рода местами для заполнения, ожидающими понятий будущих нейронаучных дисциплин. Аргументы и мотивы этих авторов были философскими, даже если принципиальное обоснование программы считалось эмпирическим.
Извинения, полученные ранними теоретиками тождества за игнорирование научных деталей, заключались в том, что нейронауки в то время были лишь зарождающейся областью, неспособной предоставить сколько-правдоподобные варианты тождеств. Однако потенциальные тождества-претенденты уже были в процессе разрабокти. Электрофизиологические демонстрации свойств рецептивного поля зрительных нейронов, проведенные Дэвидом Хьюбелом и Торстеном Визелем (Hubel and Wiesel, 1962), были опубликованы с большой помпой. Используя их методы, нейрофизиологи начали обнаруживать по всей зрительной коре нейроны, реагирующие на все более абстрактные особенности стимулов: от различения краёв до направления движения, вплоть до цветов и свойств лиц и рук. Более значимым событием, произошедшим выше десяти лет до этого, публикация Дональдом Хеббом книги «Организации поведения» (Hebb 1949). Он использует объяснения психологических явлений в терминах нейронных механизмов и анатомических схем. Объясняемые им психологические феномены включают в себя особенности восприятия, обучения, памяти и даже эмоциональных расстройств. Хебб свои объяснения в качестве вариантов тождеств. (См. Введение к его книге 1949 года). Одним из философов, действительно обратившим внимание на тот момент нейронаучные подробности, была Барбара фон Экардт Кляйн (Von Eckardt Klein, 1975). Она обсуждала теорию тождества в отношении ощущений прикосновения и давления и включила в свою работу актуальные в то время гипотезы о нейронном коде чувственной модальности, продолжительности и ощущений, которые были теоретически осмыслены Маункаслом, Либетом и Джаспером (Маунткасл, Либет и Джаспер). И все же фон Экардт Кляйн была явным исключением. Общие, доступные на тот момент нейронаучные данные игнорировались философскими приверженцами ранних теорий тождества, так и их оппонентами.
Следовательно, детальное понимание одного из реализаций физических систем (например, мозга) не проливает свет на фундаментальную природу сознания. Таким образом, психология независимой науки, изучающей какой-либо конкретный из физических реализаторов сознания (см. Статью о множественной реализуемости в SEP). Вместо нейронаук, предлагаемые услуги по анализу-ориентированные философии, обоснования и вдохновляющие идеи в когнитивной психологии и исследованиях искусственного интеллекта. Эти дисциплины абстрагируются от базовых физических механизмов и подчеркивают «информационные» свойства и способности репрезентаций (Haugeland 1985). В то же самое время, однако, нейронауки погружались в сферу познания, особенно в обучение и память. Эрик Кэндел (Кэндел 1980), к примеру, подходящие пресинаптические механизмы, регулирующие скорость высвобождения трансмиттера, в качестве цитологического объяснения простых форм ассоциативного обучения. Вместе с когнитивистскими аспектами ассоциативного обучения (например, блокирование, обусловливание второго порядка, затенение) может быть объяснены посредством клеточной биологии через комбинации этих базовых форм, реализованных в высших нейронных анатомических элементах. .
Работать над постсинаптической стороной, нейробиологи начали раскрывать клеточные долгосрочного потенцирования (Bliss and Lomo, 1973).
Психофизиологи быстро отметили объяснительный потенциал этих механизмов для различных форм обучения и памяти‹2› . И все же сравнительно немногие философы- «материалисты» обратили на это внимание. С чего бы? Большинство из них были национальными функционалистами. Они полагали, что детали «реализации уровня» («уровень реализации») могут быть важны для клинициста, но к теоретику сознания отношения не имеют.
Главным поворотным моментом в пробуждении интересаса философов к нейронаукам публикация Патриции Чёрчленд «Нейрофилософии» (Churchland 1986). Супруги Чёрчленды (Патрисия и Пол) уже известны своей пропагандой элиминативного материализма (см. Следующий раздел). В своей книге (1986) Черчленд выкристаллизовала элиминативистские аргументы предшествующего десятилетия, объединила лежащие в их основе фрагменты философии и поместила философскую прослойку между занявшим пять глав введением в нейронауки - и главой из 70 страниц, посвященной трем на тот момент теориям функционирования мозга. Она не сожалела о своих намерениях. Она представляет философию науки нейроучёным, а нейроучёным - философам. Ничто, настаивала она, не может быть более очевидным, чем релевантность эмпирических фактов о работе мозга для философии сознания. Для описания этого междисциплинарного метода она использовала термин «коэволюция» (заимствовав его из биологии). Суть этого метода - в поиске источников ресурсов для теории где угодно, не важно, находится ли источник иерархически выше или обсуждаемого вопроса. Стоя на плечах таких философов, как Куайн и Селларс, Чёрчленд настаивала: указание какой-то точки, где заканчивается нейронаука и начинается философия, безнадежно, границы плохо рассчитаны. Нейрофилософы будут выбирать ресурсы из дисциплин по своему усмотрению. как начинается указание на нее и Селларс, Чёрчленд настаивала. Нейрофилософы будут выбирать ресурсы из дисциплин по своему усмотрению. как начинается указание на нейн и Селларс, Чёрчленд настаивала: начало какой-то точки, где заканчивается наука и философия, безнадежно, как границы плохо. Нейрофилософы будут выбирать ресурсы из дисциплин по своему усмотрению.
В философском обсуждении, начатом Черчленд, преобладали три темы: разработка альтернативы теории логических эмпиристов о интертеоретической редукции; ответы на аргументы о дуализме свойств, основанные на субъективности и сенсорных квалиа; а также ответы на анти-редукционистские аргументы множественной реализуемости. Эти проекты прошли центральными в нейрофилософии свыше десятка лет после появления книги Чёрчленд. Джон Бикл (Bickle 1998) расширил основную идею Клиффорда Хукера (Hooker 1981a, b, c) о пост-эмпирической теории интертеоретической редукции.
Он количественно определил ключевые понятия, используя теоретико-модельное изложение структуры теории, адаптированное из структурной программы в философии (Balzer, Moulines, and Sneed, 1987). Он также привёл к эксплицитной форме аргумент, чтобы сделать онтологические выводы (кросс-теоретические тождества, пересмотры или исключение) из природы того, как отношения интертеоретической редукции в конкретных случаях. Например, обычно заключают, что видимый свет, - теоретическое представление из оптики, - электромагнитное излучение в пределах определенных длин волн, то есть теоретическое представление из электромагнетизма; в этом случае имеет место кросс-теоретическое онтологическое тождество. Также обычно заключают, что флогистона не существует: это - исключение типа сущностей из научной онтологии.
Пол Чёрчленд (Churchland 1996) продолжил атаку на аргументы дуальности свойств, выдвинутые в пользу нередуцируемости сознательного опыта и сенсорных качеств. Он утверждал, что получение определенных знаний в области существующей сегодня нейробиологии ощущений (сенсорная нейробиология) увеличивает способность «вообразить» или «постичь» всеобъемлющее нейробиологическое объяснение сознания. Он использует этот вывод, используя характерно задействующий воображение мысленный эксперимент, основанный на истории оптики и электромагнетизма.
Наконец, критика аргумента множественной реализуемости процветает - равно как и оспаривается - до наших дней. Хотя аргумент множественной реализуемости остается влиятельным среди нередуктивных физикалистов, ныне он потерял свое почти что универсальное признание, некогда снискал.
Появились ответы на аргумент множественной реализуемости, основанные на деталях нейронаучных исследований. Например, Уильям Бичтел и Дженнифер Мандейл (Bechtel and Mundale, 1999) утверждают, что нейроучёные используют психологические критерии в исследованиях картирования мозга. Этот факт подрывает вероятность того, что психологические сущности реализованы множественным образом (обзор последних разработок и достижений, см. В статье о множественной реализуемости в SEP).
Элиминативный материализм и нейронизированная философия
Элиминативный материализм (ЭМ) в том виде, в котором его наиболее активно пропагандируют Пол и Патриция Черчленды, является соединением двух утверждений. Во-первых, присущая нашему здравому смыслу концепция «убеждения-желания» (концепция «убеждения-желания») о характере психических событий и процессов, наша «народная психология», является ложным и вводящим в заблуждение описанием причин человеческого поведения. Во-вторых, как и другие ложные понятийные аппараты из области народных теорий и истории науки, - она будет заменена будущей нейронаукой, а не плавно редуцирована или включена в последнюю. Черчленды характеризуют народную психологию как набор проповедей общего толка, призванного. Вы спрашиваете, почему Марика не сопровождает меня этим вечером. Я отвечаю, что нашему внуку нужна была сиделка. Вы сочувственно киваете. Вы понимаете мое объяснение. Это - всего лишь одна из огромного количества проповедей о причинах человеческих нарушений, которые, по утверждению ЭМ, столь велики, что не подлежат пересмотру. Хотя приведённый пример включает в себя только убеждения и желания, народная психология обладает возможностями репертуаром пропозициональных установок в объяснительной обойме: надежды, намерения, страхи, фантазии и многое другое. Сторонники ЭМ предсказывают, что в будущем по-настоящему научная психология или нейронауки в конечном итоге откажутся от всего этого словаря и заменят егоми,
ЭМ является физикалистской программой в полностью традиционном философском смысле. Он постулирует, что некоторая будущая наука о мозге в Правиле предоставит правильный отчёт о (человеческом) поведении. Этот материализм является элиминативным в том плане, что предсказывает будущее изгнание народно-психологических типов типов нашей пост-нейронаучной онтологии. Сторонники ЭМ часто используют научные аналогии (Фейерабенд, 1963; Пол Черчленд, 1981).
Окислительные сходные реакции, описанные в элементарной химии, не имеют значения с высвобождением флогистона. Даже «направление», когда объект горит (или ржавеет), связан, флогистон же считался улетучившимся. Результатом этого теоретического изменения стало исключение флогистона из нашей научной онтологии. Там больше нет такие вещи. Согласно ЭМ, по тем же причинам продолжающимся развитие в нейронауках покажет, что не существует таких вещей, как убеждения, желания и остальные пропозициональные установки, - в виде, как их характеризует здравый смысл.
В данной статье мы сосредоточены лишь на том, как результаты нейронаучных исследований придали окончательный вид аргументам в поддержке ЭМ. Удивительно, но особенно сильное влияние на себе испытал лишь один аргумент. (Основные аргументы в пользу ЭМ подчеркивают неудачи народной психологии в качестве объяснительной теории поведения.) Этот аргумент основан на достижении в когнитивной и вычислительной нейробиологии, способном обеспечить подлинную альтернативу репрезентациям и вычислениям, подразумеваемым в народно-психологических обобщениях. Многие элиминативные материалисты предполагают, что народная психология ориентирована на пропозициональные репрезентации и вычисления на основе их содержаний, которые имитируют бытовые умозаключения‹3›(Пол Черчленд, 1981; Стич, 1983; Патрисия Черчленд, 1986). [2]
Некоторые элиминативисты считают, что нейробиология начала предлагать такого альтернативу только в последние тридцать лет. Ключевой характерной чертой этой альтернативы использование точек уровня событий в пространстве в интерпретации на синаптических и паттернов нейронных активностей в биологических и искусственных нейронных сетях. Различия между понятиями когнитивной репрезентации и преобразований, осмысленными векторно-пространственной точкой зрения с одной стороны, и с тем, как ониаются в свете пропозициональных установок, народной психологии с другим, основанием для одного аргумента в пользу ЭМ (Пол Черчленд, 1987). Тем не менее, суть этого аргумента будет непрозрачна для тех, кто не осведомлён в когнитивной и вычислительной нейробиологии, поэтому мы представим некоторые детали. Разобравшись с этими средствами, мы вернемся к опирающимся на них аргументам в пользу ЭМ (пятью абзацами далее).
На уровне базового вычислительного элемента нейронной сети, биологической или искусственной, является нервная клетка или нейрон. Математически представлены простые вычислительные устройства, преобразующие входные данные в выходные. И входы, и выходы отражают биологические переменные.
Для нашего обсуждения предположим, что нейрональные входы - это частота потенциалов действия (нейронные «спайки») в аксонах, чьи оконечные ветви (концевые ветви) соединяются посредством синапса с рассматриваемым нейроном, а нейрональный выход - это частота потенциалов действия, генерируемых в аксоне данного нейрона после обработки входных сигналов. Таким образом, нейрон вычисляет свой общий вход, математически обычно рассматриваемый как сумма произведений силы сигнала на каждой линии входа, умноженной на синаптический вес‹4› на этой линии. Затем нейрон вычисляет новое состояние активации на основе своего общего входа и текущего состояния активации, а также новое состояние активации на основе своего нового значения активации. Выходное состояние нейрона передается как мощность сигнала в направлении любого из нейронов, с его аксон соединяется посредством синапсов. Выходное состояние упорядоченно отражает новое состояние активации нейрона.‹5›
Рассмотренные таким образом, биологические и искусственные нейронные сети естественным образом интерпретируются как векторные-данные преобразователи.
Входной вектор состоит из значений, отражающих паттерны активности в аксонах, соединяющихся посредством синапсов с сетью нейронов извне (например, от сенсорных преобразователей или других нейронных сетей).
Выходной вектор состоит из значений, отражающих генерируемые в нейронах сети паттерны активности, которые передаются за пределы сети (например, на моторные эффекторы или другие нейронные сети). Учитывая, что активность каждого нейрона отчасти зависит от их входа, отчасти зависит от синаптических весов (например, скорости высвобождения пресинаптических нейротрансмиттеров, количества и эффективности рецепторов постсинаптических нейротрансмиттеров, наличия энзимов в синаптической щели), способности биологических сетей к изменению их синаптических весов делает их пластичными векторно-векторными преобразователями.
Принципиально, биологическая сеть с пластичными синапсами может реализовать любое векторное преобразование, которое позволяет составить (количество входных единиц, выходных единиц, уровней обработки, возвратности, перекрестных связей и т.д.) (обсуждается в книге Пола Чёрчленда (Churchland 1987)) со ссылками на первичную научную литературу)).
Рис 1.
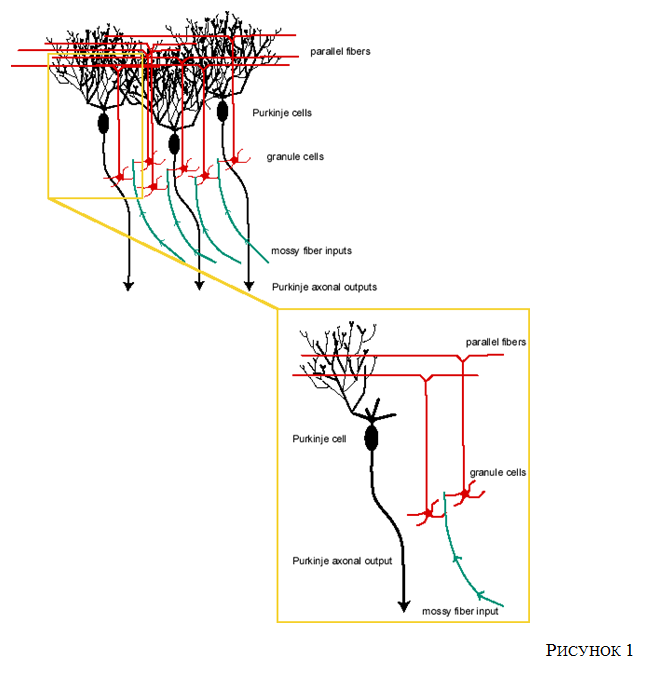
Анатомическая организация мозжечка представляет собой непосредственный пример сети, поддающейся приведенной выше вычислительной интерпретации.
Рассмотрим Рисунок 1.
Мозжечок представляет собой выпуклую извилистую структуру, дорсальную‹6› к стволу мозга. Разнообразные исследования (поведенческие, нейропсихологические, проводимые на отдельных нейронах, электрофизиологические) указывают на вовлеченность этой структуры в двигательную интеграцию и тонкую моторную координацию.
Мшистые волокна‹7› (аксоны [см. в нижней части рисунка, они задают вход - вход мшистых волокон]), идущие из нейронов вне мозжечка соединяются посредством синапсов с мозжечковыми гепариноцитами [упомянутые в сноске «клетки-зёрна», гранулярные клетки], которые, в свою очередь, передают сигнал параллельным волокнам [параллельные волокна]. Паттерны активности в общей массе мшистых волокон (частота потенциалов действия на единицу времени в каждом волокне, передаваемом в мозжечок) значения для входного события. Параллельные образуют множественные синаптические связи с дендритными деревьями и клеточными мозжечковыми нейронами Пуркинье [нейроны Пуркинье / клетки Пуркинье]. Каждый нейрон Пуркинье «суммирует» свои постсинаптические потенциалы (ПСП) и испускает последовательность потенциалов действия вниз по своему аксону, исходя (частично) из своего общего входа и предыдущего состояния активации. Аксоны Пуркинье передают сигнал за пределы мозжечка [вывод аксонов Пуркинье]. Таким образом, выходной вектор сети представляет собой упорядоченные значения, представляющие паттерны активности, сгенерированные в каждом аксоне Пуркинье. Изменения в эффективности отдельных синапсов параллельных волокон и нейронов Пуркинье, в свою очередь, приводят к иным итоговым величинам ПСП в аксонах Пуркинье, генерируя в результате другие частоты аксональных спайков. В вычислительном отношении это соответствует другому выходному вектору при том же паттерне входной активности - это и есть представляющие паттерны активности, сгенерированные в каждом аксоне Пуркинье. Изменения в эффективности отдельных синапсов параллельных волокон и нейронов Пуркинье, в свою очередь, приводят к иным итоговым величинам ПСП в аксонах Пуркинье, генерируя в результате других частот аксональных спайков. В вычислительном отношении это соответствует другому выходному вектору при том же паттерне входной активности - это и есть представляющие паттерны активности, сгенерированные в каждом аксоне Пуркинье. Изменения в эффективности отдельных синапсов параллельных волокон и нейронов Пуркинье, в свою очередь, приводят к иным итоговым величинам ПСП в аксонах Пуркинье, генерируя в результате других частот аксональных спайков. В вычислительном отношении это соответствует другому выходному вектору при том же паттерне входной активности - это и естьпластичность‹8› .
Рис 2.
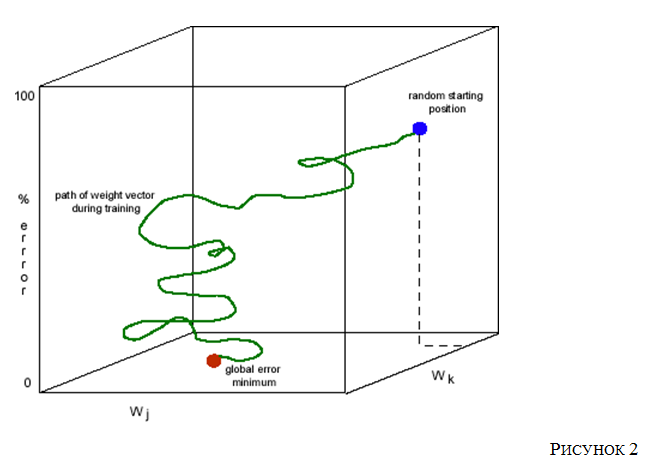
Приведённая интерпретация делает весьма полезные математические ресурсы теории динамических систем доступными для использования вычислительными нейробиологами: примером служат пространства . С опорой на них процесс обучения можно плодотворно охарактеризовать в терминах изменения синаптических весов в сети и последующего уменьшения ошибок на выходе сети. (Этот подход к обучению восходит к работе ( Hebb 1949), хотя векторно-пространственная интерпретация не была частью описания Хебба.) Чтобы удобнее представить себе предложенное описание, используется пространство синаптического веса-веса.. Одно из его измерений представляет собой глобальные ошибки для данной задачи на выходе. Рассмотрим Рисунок 2 . Точки в этом многомерном пространстве состояний представляют глобальную ошибку выполнения ( глобальная производительность ошибка ), связанный с каждым из наборов синаптических весов в сети. В соответствии с биологическим алгоритмом обучения, веса меняются с каждым выполнением, глобальная ошибка выполнения сети постоянно уменьшается. С каждым эпизодом обучения изменению синаптических весов сети увеличивает общую ошибку выходного вектора сети, рассчитанную по сравнению с требуемым выходнымом для входного вектора. Тем самым обучение как изменение синаптических весов, скоррелированное с понижением по оси в ошибки (Churchland, Sejnowski, 1992). Представления (понятия) можно изобразить в виде разбиений в многомерных векторных пространствх; одним из примеров этого цифрового пространства активации нейронов .
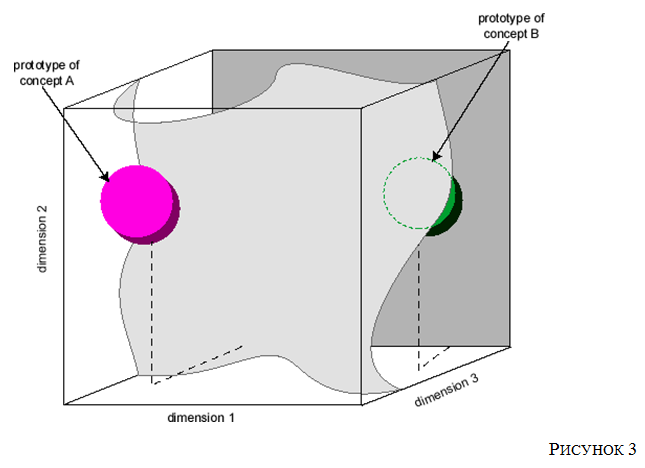
Взгляните на Рисунок 3 .
График такого рода включает в себя по одному измерению для значений активации каждого нейрона в сети (или некоторого определенного пространства подмножества нейронов сети, - например, находящихся в определенном слое). Точка в этом многомерном пространстве представляет собой один из агентов паттернов всех нейронов в сети. Сгенерированные входными объектами вектора паттерны активности, сеть, которая научилась группировать, будут образовывать кластер вокруг (гипер-) точки или частичного объема в векторном пространстве активности. Иными словами: любой другой паттерн, элемент подобной этой группы, создаст объект взаимодействия, лежащий в геометрической точке этой точки или частичному объему. Пол Чёрчленд ( Черчленд 1989 г.) утверждал, что такая такая теория формальной психологии наконец-то использует основу для вдохновлённой нейронной моделями основы для прототипических теорий понятий, разработанных в когнитивной психологии конца ХХ века.
Рис 3.
Используя это теоретическое достижение уже в области нейрофилософии, Пол Чёрчленд (Черчленд, 1987, 1989 г.) использует новый, вдохновлённый нейронаучными исследованиями аргумент в пользу ЭМ. Согласно только что изложенной интерпретации работы нейронных сетей, простейший вид преобразования вычислений в мозге. Эта перспектива резко контрастирует с пропозициональными репрезентациями и логическими / семантическими вычислениями, постулируемыми народной психологией. Содержание векторов, - упорядоченная последовательность действительных чисел, - незнакомо и чуждо здравому смыслу. Это кросс-теоретическое концептуальное различие как минимум столь же велико, как понятия окислителя и флогистона, или понятиями тепла в молекулярно-кинетической теории и в теории теплородного флиюда. Флогистон и теплородный флиюд - два «показательных» примера типов сущностей, исключительных из нашей научной онтологии в силу характера интертеоретических связей между теориями, которые они (эти сущности) соединены, и теориями, которые пришли им на смену. Структурные и динамические различия между народно-психологическими и последвшими в то время когнитивно-нейробиологическими типами сущностей давали основание предположить, что теории, связанные с ними, схожим образом заменят теорию, связанную с первыми. Однако это утверждение, бывшее ключевой целевой элиминативистского аргумента, само базировалось на предсказываемых интертеоретических отношениях. А с распространением нейронных сетей и параллельной распределенной обработки интертеоретические контрасты с объяснительными типами сущностей, предлагаемыми народной психологией, перестали быть просто надеждой элиминативистов на будущее. Вычислительная и когнитивная нейробиология наконец-то было представлено альтернативную кинематику для процессов познания - причём такую, у которой не структурных аналогов пропозициональным установкам народной психологии или логико-подобным вычислением над пропозициональными содержаниями.
Безусловно, векторно-пространственные альтернативы приведённой интерпретации работы нейронных сетей чужды народной психологии. Но оправдывают ли они ЭМ? Даже если пропозициональное содержание народно-психологических положений не находит аналогов в одном теоретическом достижении из области когнитивной и вычислительной нейробиологии (это было острой темой три десятилетия назад), могут существовать и другие аспекты познания, в народной психологии верна. В рамках научного реализма, придавшего форму ранней нейрофилософии, делался следующий вывод: истинно ли утверждение о кросс-теоретической идентичности (например, описываемое народной психологией состояние F идентично нейронному состоянию N), или же истинно утверждение элиминативиста (не существует такой вещи, как описываемое народной психологией состояние F) - это зависит от характера интертеоретической редукции между теориями, связанными рассматриваемыми утверждениями (Hooker 1981a, b, c; Churchland 1986; Bickle 1998). Но подразумеваемый подход интертеоретической редукции также широкий спектр редукций, начиная от «гладких» до «весьма ревизионных»«Край турбулентных»‹9› .
Может ли редукция народной психологии к «векторной» вычислительной нейробиологии занять какое-то среднее положение между «гладкими» и «турбулентными» концами интертеоретической редукции и, следовательно, предложение «ревизионный» вывод? Потенциальной аналогией в данном случае может служить редукция классической равновесной термодинамики к статистической механике. Исходя из эмпирических данных, Джон Бикл (Бикл 1992, 1998, глава 6) утверждал, что такой исход вполне вероятен. Определены условия «ревизионных» редукций из различных примеров и примеров использования сложных механизмов между народной психологией и когнитивной нейробиологией по мере развития последней. В частности, народная психология, по-видимому, правильно поняла небрежно очерченные функциональные профили многих когнитивных состояний, - особенно тех, которые связаны с сенсорными входами и поведенческими выходами. Похоже, что она правильно понимает «интенциональность» многих когнитивных состояний (объект, из которого состоит это состояние или которое на котором установлено), пусть даже когнитивная нейробиология и избегает своего неявного лингвистического объяснения этой особенности. Ревизионный физикализм предсказывает основные концептуальные изменения народно-психологических понятий, но отрицает полное уничтожение разнообразия по типу теплорода-флогистона.
В предисловии к своей книге «Нейрокомпьютерная перспектива» (Churchland 1989) Пол Чёрчленд подчёркнул нейрофилософски утверждал, что в скором времени станет невозможным провести серьезную работу в философии науки, не опираясь на эмпирическую работу в области мозга и поведенческих наук. Чтобы оправдать это утверждение, во второй части книги он нейрокомпьютерные переформулировки ключевых понятий философии науки. В основе его переформулировок лежит нейрокомпьютерный подход к структуре научных теорий (1989: гл. 9). Проблемы с ортодоксальным взглядом на научные теории как «наборы предложений» были хорошо известны с 1960-х годов. Чёрчлендал за замену ортодоксального взгляда на перспективу, вдохновленную «векторной» интерпретацией активности нейронной сети. Реализованные в нейронных сетях (как было показано выше) представленные системы, соответствующие значимым различиям во внешней среде, но сами явным образом не представлены во входной совокупности (входной корпус) и разрешенной сети реагировать на входные данные так, чтобы уменьшать ошибку . Согласно Черчленду, в этом и состоят функции теорий. Чёрчленд смело утверждал: чья-то конкретная теория мира - это особая точка в векторном пространстве синаптического веса-ошибки данного индивида.
Именно конфигурация синаптических весов разбивает активационное пространство индивидуума на меньшие сегменты, тем самым большее число будущих сообщений об ошибках как для уже знакомых, так и для новых входных данных. (Вновь обрати к Рисунку 2 и Рисунку 3). Впрочем, предложенная переформулировка возражения. Чёрчленд гордится тем. Но, как отметил Бикл (Bickle 1993), нейрокомпьютерные модели, основанные на выше математических средствах, далеко продвинулись в области математической абстракции. Они представляют собой не что, как новое (хотя и наводящее на размышления) применение математики квазилинейных динамических систем к упрощенным схемам межнейронных связей в мозге. Нейрофилософы могут предоставить сколько-нибудь внятное описание отождествлений в онтологических категориях (репрезентации векторов и их преобразование - но во что именно?), Прежде чем сообщество философов, начнёт рассматривать теории как точки в многомерных пространствах состояний, реализованные в биологических нейронных сетях. (Отметим, что в возражении Бикла скрывается важное методологическое допущение, мы обсудим в конце следующего абзаца.) Нейрофилософы могут предоставить сколько-нибудь внятное описание отождествлений в онтологических категориях (репрезентации векторов и их преобразование - но во что именно?), Прежде чем сообщество философов, начнёт рассматривать теории как точки в многомерных пространствах состояний, реализованные в биологических нейронных сетях. (Отметим, что в возражении Бикла скрывается важное методологическое допущение, мы обсудим в конце следующего абзаца.) Нейрофилософы могут предоставить сколько-нибудь внятное описание отождествлений в онтологических категориях (репрезентации векторов и их преобразование - но во что именно?). (Отметим, что в возражении Бикла скрывается важное методологическое допущение, мы обсудим в конце следующего абзаца.)
Нейрокомпьютерная переформулировка Черчлендом используемых научных и эпистемологических концепций опирается на описанный выше подход к теориям. Он делает наброски «нейронизированных» описаний для такой тем философии, как теоретическая нагруженность восприятия, природа объединения понятий, достоинство теоретической простоты, природа парадигм Куна, кинематика концептуальных изменений, характер абдукции, природа объяснения и даже нравственное знание вместе с эпистемологической нормативностью. Пример, концептуальная реорганизация - активация уже существующего прототипа области разбиения многомерного пространства в обученной нейронной сети с помощью входного паттерна нового типа. Очевидно, Мы не сможем изучить предложения в их первоначальном виде. Мы не можем предоставить должное многочисленным и разнообразным попыткам Чёрчленда по нейрофилософскому переформуыванию тем философии: заинтригованного читателя. Однако сюда напрашивается ремарка о философской методологии автора. Чёрчленд явно не делает попытку «концептуального анализа» ни в какой форме, напоминавшей бы последний в традиционном философском смысле. Как правило, этого не делают ни в одном из своих проектов переформулировывания и другие нейрофилософы. (Вот почему обсуждение нейрофилософских переформулировок совпадает с обсуждением ЭМ.) Есть философы, которые обеспечивают идеальным анализом дисциплины относительно простой набор необходимых и достаточных условий, выраженных на не-техническом естественном языке и регулирующих применение значимых понятий (например, справедливости, знания, теории или объяснения). Насколько это возможно, подобные анализы должны согласовываться с до-теоретическим использованием. В идеале, они вообще должны быть синонимию. Другие философы рассматривают такой идеал анализа как бесплодный, крайне ошибочный в отношении глубинной структуры человеческие знания (Ramsey 1992). Нейрофилософы, как правило, располагаются в последней группе. Те, кому не нравятся философские рассуждения насчёт и возможности развития науки переформулировать «(пе-реформами-ул-ировать») традиционные философские концепции, вероятно, уже представлены, что нейрофилософия - не для них. Однако знакомое обвинение в том,
Прежде, чем мы оставим тему предлагаемых нейрофилософских применений теоретической разработки (сделанной на основе когнитивной / вычислительной нейробиологии в стиле «нейронных сетей»), следует сделать ещё один, заключительный пункт насчёт реальных деталей. Взятый Чёрчлендом на вооружение подход не задержался на переднем крае вычислительной нейробиологии надолго. Многие разработчики моделей нейронов вскоре отказались от этого подхода к моделированию мозга. Компартмент (почастное) моделирование вычислительным нейробиологам имитировать активную активность между отдельными участками нейронной мембраны (Bower and Beeman 1995). В свою очередь, этот подход позволил разработчикам моделей контролировать и управлять различными внутриклеточными факторами, определяющими потенциалы действия в единицу времени,
К середине 1990-х годов разработчики моделей быстро начали калибровать нейроны в своих целевых цепях (целевые схемы) «по заказу». Все более мощное компьютерное оборудование по-прежнему им изучать характеристики цепи моделируемых сетей.
В силу этих факторов многие серьезные вычислительные нейробиологи переключились на работу на уровень анализа, рассматривающий нейроны как структурированные, нежели простые вычислительные устройства. Благодаря компартментному моделированию вектор-системы оказались менее полезны в серьезных нейробиологических моделях: они были заменены дифференциальными уравнениями, описывающими ионные токи через участки мембраны нейрона. В полученных моделях оказалось больше биологических деталей, чем позволяли получить «коннекционистские» модели. Это методологическое изменение в вычислительной нейробиологии означало, что опирается на «коннекционистские» ресурсы нейрофилософия перестала соответствовать современному уровню науки.
Философия науки и научная эпистемология были не единственными областями, в которых нейрофилософы педалировали нейробиологических открытий для традиционных философских тем. Десять лет спустя после публикации «Нейрофилософии», Кэтлин Экинз (Akins 1996) взялась утверждать, что в основе разнообразных сложных «натуралистических» программ о природе интенциональности лежит «традиционный» взгляд на чувства. (В качестве примеров она приводит Черчлендов, Даниэля Деннетта, Фреда Дрецке, Джерри Фодора, Дэвида Папино, Денниса Стампе и Кима Стерли.)
Недавняя - на момент дискуссии - нейронаучная работа о механизмах и стратегиях кодирования, реализованных в сенсорных рецепторах, демонстрирует ошибочность традиционного взгляда. Традиционное мнение считает, что сенсорные системы являются «достоверными», по крайней мере, на 3-х основаниях:
- (1) Каждый сигнал в системе коррелирует со сравнительно небольшим диапазоном свойств внешней (для организма) среды.
- (2) Структура внешних отношений, к которым чувствительны рецепторы, сохраняющие сенсорные структуры внутренних отношений между соответствующими состояниями.
- И (3) сенсорная система правдиво, без вымышленных дополнений или приукрашиваний, реконструирует внешние события.
Используя в качестве недавних на тот момент нейробиологические открытия о свойствах отклика тепловых устройств в коже (т.е. «терморецепторов»), Экинс показала, что сенсорные системы скорее «нарциссические», нежели «достоверными». Все три допущения нарушаются.
Эти нейробиологические детали и их философские импликации ставят новые вопросы перед философией восприятия и основами натуралистических проектов интенциональности.
Взяв на вооружение изученную нейрофизиологию сенсорных рецепторов, нашу «философию восприятия» или подход к «перцептивной интенциональности» больше не будет фокусироваться на поиске корреляций между состояниями сенсорных систем и «достоверно обнаруженными» внешними свойствами. Этот философский (и научный!) Проект основывается на ошибочном понимании чувственного восприятия как «достоверного».
Нейронаучные знания о деятельности сенсорных рецепторов также показывают, что сенсорный опыт не служит натуралисту и в качестве «просто парадигмального случая» интенционального отношения между репрезентацией и миром. В который раз, ставшие доступными научные детали показали наивность некоторых философских проектов.
Сосредоточив внимание на анатомии и физиологии системы передачи боли, Валери Хардкасл (Hardcastle 1997) подтвердила аналогичные отрицательные выводы и для другого популярного методологического допущения. Они использовали для анализа и теоретизирования о сознательном опыте в целом.
Тем не менее, точка зрения о переживаниях была вполне обоснована: элиминативизм, различные объективистские взгляды, реляционные подходы, а также субъективные перспективы.
Почему же, без возможности использования в том, что именно с болевых переживаний можно начинать анализ или теорию сознания, между позициями так мало согласия? Хардкасл призвала рассмотреть два ответа.
Во-первых, как правило, философы не осведомлены о нейрональной сложности наших систем передачи боли и строят свои анализы или теории на результирующем действии одного компонента многокомпонентной системы.
Во-вторых, даже те, кто понимает некоторые из основ нейробиологии, склонны защищать теории воротного контроля‹10› .
Однако даже лучший из способов воротного контроля расплывчаты в отношении нейронных механизмов действия «ворот».
Вместо этого Хардкасл предложила раздельно-двойную систему передачи боли, которая из своей сенсорной системы боли, - которая по своему нейробиологическому применению другой сенсорной системы боли, - и нисходящей системы подавления боли. Она утверждает, что эта двойственная система согласуется с нейробиологическими открытиями и учитывает все болевые явления, искушающие философов строить (но ограниченные) теории болевого опыта. Нейробиологическая уникальная система подавления боли, контрастирующая с механизмом других сенсорных модальностей, делает обработку боли нетипичной.
В частности, система подавления отделяет болевые ощущения от стимуляции ноцицепторов (болевых рецепторов).
Нейронаука и психосемантика
Развитие и обоснование содержания содержания занимает центральное место в современной философии сознания. Объектом общих устремлений в дискуссиях на эту тему является теория когнитивной репрезентации, согласующаяся с физической или натуралистической онтологией. Здесь мы опишем несколько значимых вкладов, которые в этот проект внесла нейрофилософия.
Когда человек воспринимает или вспоминает, состояние его мозга обладает интенциональностью или «направленностью на» («направленностью на»). Восприятие или воспоминание направлено на то, что у человека закончилось кофе; оно репрезентирует нехватку кофе у человека. Репрезентативное (представительное) состояние имеет содержание.
Физикалистская психосемантика стремится сделать это, используя исключительно физические науки. Нейрофилософы внесли свой вклад в два типа физикалистских психосемантик: функционально-ролевой подход и информационный подход. С описанием этих и других теорий ментального содержания можно познакомиться с SEP о причинных теориях ментального контента , ментальной репрезентации и телеологических теориях ментального контента.
Основное утверждение семантики функциональных ролей заключается в том, что репрезентация обладает своим конкретным содержанием в силу отношений, которые она поддерживает с другими репрезентациями. Парадигмальным примером её применения являются истинностно-функциональные логические понятия, такие как конъюнктивное «и» или дизъюнктивное «или». Физическое событие воплощает собой экземпляр функции «и» в том случае, если оно отображает два истинностных входа и один истинный выход. Вот почему именно отношения, которые одно выражение устанавливает с другими, придают ему смысловое содержание «и». Сторонники функционально-ролевой семантики предлагают аналогичные анализы для содержания любых репрезентаций (Block 1995). Физическое событие является репрезентацией, к примеру, птиц, если оно имеет правильное отношение к событиям, представляющим перья, и тем, которые представляют клювы.
В отличие от этого подхода, информационная семантика приписывает состоянию содержание в зависимости от причинно-следственных связей, возникающих между состоянием и объектом, который оно представляет. Физическое состояние представляет, к примеру, птиц, в том случае, если между ним и птицами возникает подобающая причинная связь. В основе информационной семантики лежит причинный подход к информации (Dretske 1981, 1988). Красные пятна на лице несут информацию о том, что у человека корь, потому что вирус кори – причина красных пятен.
Распространенная критика информационной семантики состоит в том, что простой причинной ковариации (сопряжённого изменения) для репрезентации недостаточно, поскольку информация (в причинном смысле) по определению всегда достоверна, в то время как репрезентации могут искажать данные.
Популярное решение этой проблемы включает в себя телеологический анализ «функции». Состояние мозга представляет X благодаря функции передачи информации о том, что X является его причиной (Dretske 1988). Этими двумя подходами популярные варианты психосемантики не исчерпываются, но именно в них нейрофилософы вносят наибольший вклад.
Приверженность Пола Черчленда семантике функциональных ролей восходит к его самым ранним взглядам на семантику терминов в языке. В своей книге (Churchland 1979) он настаивал на том, что семантическая идентичность (содержание) термина может быть выведено из места последнего в сети предложений всего языка. Предполагаемые ранними сторонниками семантики функциональных ролей функциональные структуры были сетями, чьи узлы соответствуют объектам и свойствам, обозначенным выражениями в языке. Таким образом, имеющий подобающие связи узел может представлять птиц, другие перья и другие клювы. Активация одного из узлов будет распространяться на другие. По мере развития «коннекционистского» нейросетевого моделирования (как обсуждалось в предыдущем разделе выше) у этого «локалистского» подхода с одним представлением на один узел появились альтернативы. К тому времени, когда Черчленд (Churchland 1989) представил тщательную нейронаучную проработку семантики функциональных ролей для когнитивных репрезентаций в целом, он и сам отказался от «локалистской» интерпретации. Вместо этого он предложил «семантику пространства состояний».
В предыдущем разделе мы видели, как (векторные) пространства состояний обеспечивают интерпретацию паттернов активности в нейронных сетях: как биологических, так и искусственных. Семантика пространства состояний для когнитивных репрезентаций является разновидностью семантики функциональных ролей, поскольку сохраняется общая идея: индивидуация конкретного состояния зависит от отношений, возникающих между ним и другими состояниями. Репрезентация – это точка в соответствующем пространстве состояний, а точки (или частичные объёмы) в пространстве индивидуируются своими отношениями с другими точками (местоположениями, в смысле геометрической близости).
Пол Черчленд (Churchland 1989, 1995) проиллюстрировал семантику пространства состояний для нейронных состояний, обратившись к сенсорным системам. Одной из популярных теорий сенсорной нейробиологии о том, как мозг кодирует сенсорные качества (например, цвет), является подход противоположного процесса (Hardin 1988). Черчленд (Churchland 1995) описывает трехмерное активационное векторное пространство состояний, в котором каждый воспринимаемый человеком цвет представляется в виде точки (или частичного объёма).
Каждое измерение соответствует уровням активности в одном из трех классов фоторецепторов, присутствующих в сетчатке человека, и их эфферентных путях: красно-зеленый оппонентный путь, желто-синий оппонентный путь и черно-белый (контрастный) оппонентный путь.
Попадающие на сетчатку фотоны преобразуются фоторецепторами, вызывая соответствующую степень активности в каждом из выделенных выше путей. Следовательно, репрезентация цвета представляет собой триплет значений частот активации нейронов.
В качестве иллюстрации вновь обратимся к Рисунку 3. Каждое измерение в этом трехмерном пространстве будет представлять среднюю частоту потенциалов действия в аксонах одного из классов ганглионарных клеток, выступающих из сетчатки. Каждый воспринимаемый людьми цвет будет некоторой областью такого пространства. Например, оранжевый стимул вызывает относительно низкий уровень активности как в красно-зеленом, так и в желто-синем оппонетных путях (по осям x и y соответственно), и среднюю активность в черно-белом (контрастном) оппонетном пути (ось z). С другой стороны, розовые стимулы вызывают низкую активность в красно-зеленом оппонетном пути, среднюю активность в желто-синем оппонетном пути и высокую активность в черно-белом (контрастном) оппонетном пути‹11›.
Расположение каждого цвета в пространстве создает «цветовое тело». Расположение на этом теле, а также геометрическая близость между местоположениями отражают структурные сходства между воспринимаемыми цветами. Вкусовые репрезентации человека - это точки в четырехмерном пространстве состояний, каждое из измерений которого кодирует степень активности, генерируемую вкусовыми стимулами в каждом из типов вкусового рецептора (сладкий, соленый, кислый и горький) и их отдельных эфферентных путях. Будучи реализованным в нейросети со структурными ресурсами – и, следовательно, вычислительными ресурсами, – столь же обширными, какими обладает человеческий мозг, психосемантический подход пространства состояний порождает теорию содержания для огромного числа когнитивных состояний‹12›.
В работе Джерри Фодора и Эрнеста Лепора (Fodor and LePore 1992) был сформулирован весьма сильный вызов психосемантике Чёрчленда. Сама по себе, схема с местоположением в пространстве состояний кажется недостаточной, чтобы решить проблему репрезентативного содержания состояния. Черчленд никогда не объясняет, почему точка в трехмерном пространстве состояний представляет именно цвет, нежели любое другое качество, объект или событие, которые изменяются по трем измерениям‹13› . Соответственно, подход Чёрчленда достигает своей объяснительной силы благодаря интерпретации, которую получают измерения (пространства состояний). Фодор и Лепор утверждали, что Черчленд никогда не определял, каким образом измерение представляет, например, степень солености, а не желто-синюю оппоненту длины волны. Один очевидный ответ апеллирует к стимулам, формирующим «внешние» входы в рассматриваемую нейронную сеть. Тогда, например, условия индивидуации нейронных репрезентаций цветов состоят в том, что оппонетные обрабатывающие нейроны получают входные данные от определенного класса фоторецепторов.
В свою очередь, активирующими стимулами последних выступает электромагнитное излучение (определенной части видимого спектра). Однако подобное обращение к «внешним» стимулам как к конечным условиям индивидуации репрезентируемого содержания делает полученный подход версией информационной семантики. Согласуется ли этот подход с другими нейробиологическими данными?
С нейробиологической точки зрения, парадигмальным элементом информационной семантики является детектор признаков: это – один или несколько нейронов,
- (i) максимально чувствительных к определенному типу стимулов и
- (ii) имеющих функцию указывать на наличие стимула данного типа. Примерами таких типов стимулов для детекторов визуальных признаков являются высококонтрастные края, направление движения и цвета. Излюбленным среди философов детектором признаков является предполагаемый детектор мух, которым обладает лягушка. Леттвин и соавт. (Lettvin et al. 1959) идентифицировали в сетчатке лягушки клетки с максимальной реакцией на небольшие формы, движущиеся через поле зрения
Идея, что активность этих клеток имела функцией обнаружение мух, основывалась на сведениях о питании лягушек. (в работе Bechtel 1998 представлена содержательная дискуссию.) Используя экспериментальные методы, – от регистрации отдельных клеток до изощрённой функциональной визуализации, – нейробиологи обнаружили множество нейронов, которые максимально чувствительны ко множеству сложных стимулов. Однако установить условие
- (ii) для детектора признаков намного сложнее. Даже некоторые парадигмальные примеры были поставлены под сомнение. Удостоенную Нобелевской премии Работу Дэвида Хьюбела и Торстена Визела (Hubel and Wiesel1962) по определению рецептивных полей нейронов в первичной (зрительной) коре часто интерпретируют как выявление клеток, чьей функцией является обнаружение краев. Однако Лехки и Сейновски (Lehky и Sejnowski 1988) оспорили эту интерпретацию. Они обучили искусственную нейронную сеть отличать трехмерную форму и ориентацию объекта в пространстве от его двумерного паттерна затенения. Их сеть включает в себя множество функций визуальной нейрофизиологии. Узлы в обученной сети оказались максимально чувствительными к контрастам краев, но, по-видимому, не имели функции обнаружения краев. (См. обзор в Churchland and Sejnowski 1992)
Кэтлин Акинс (Akins 1996) предложила нейрофилософский вызов другого рода для информационной семантики и связанного с ней представления о сенсорной репрезентации, опирающегося на обнаружение признаков. В предыдущем разделе мы познакомились с утверждениями Акинс, что физиология терморецепции нарушает три необходимых условия «достоверной» репрезентации. Исходя из этого, она поставила под сомнение необходимость искать нейроны-детекторы признаков, чтобы обосновать психосемантику в целом, – включая содержание мыслей. Например, человеческие мысли о мухах чувствительны к численным различиям между конкретными мухами и конкретными местами, которые те могут занимать. Но цели, сводящиеся к питанию лягушки, хорошо удовлетворяются и без репрезентативной системы, чувствительной к таким онтологическим тонкостям. В знании о том, является ли замеченная сейчас муха численно идентичной мухе, замеченной минуту назад, нет необходимости – а, вероятно, оно может и вовсе не фигурировать в репертуаре обнаруживаемых лягушкой признаков. Критика Акинс заставляет усомниться, могут ли детали сенсорных преобразований быть масштабно увеличены, чтобы обеспечить адекватно унифицированную психосемантику для всех понятий. Её¬ работы также подняли новые вопросы относительно человеческой интенциональности. Как мы вообще переходим от паттернов активности в «нарциссических» сенсорных рецепторах (привязанных не к «объективным» особенностям окружающей среды, а, скорее, только к воздействию стимулов на возбуждённый участок ткани) к человеческим онтологиям, изобилующим устойчивыми объектами со стабильными конфигурациями свойств и отношений, типов и их знаков (как показывает приведенный выше пример «мыслей о мухах»), и всем остальным? И каким образом развитие стабильной, богатой онтологии дало преимущества для выживания предкам людей?
Объяснённое сознание?
За последние три десятилетия сознание вновь вышло на передний план в качестве темы исследований в философии сознания, а также в когнитивных науках и науках о мозге. Вместо того, чтобы игнорировать разум, многие физикалисты попытались его объяснить (Dennett 1991). В данном разделе мы сосредоточимся исключительно на том, как нейробиологические открытия повлияли на философские дебаты о природе сознания и его связи с физическими механизмами. (См. ссылки на другие записи в SEP в связанных статьях: в них представлена более широкая панорама дискуссий о сознании и физикализме.)
Широко известен аргумент Томаса Нагеля (Нагель 2003) о том, что сознательный опыт субъективен и, следовательно, постоянно сопротивляется объективно-научному способу понимания. Нагель предложил нам задуматься над тем, «каково быть летучей мышью», и обратился к интуитивному суждению, что никакой объём физико-научных знаний, - включая нейробиологические, - не даст полного ответа. Введённые Нагелем в оборот интуиции вызвали широкую философскую дискуссию. По меньшей мере два широко известных ответа напрямую касались нейрофизиологии. Джон Биро (Biro 1991) предположил, что часть введённой Нагелем в оборот интуиции, будто опыт летучих мышей существенно отличается от опыта человека, сама предполагает систематические отношения между физиологией и феноменологией. Кэтлин Акинс (Akins 1993a) углубилась в изучение существующих знаний о физиологии летучих мышей и многое сообщила касательно вопроса Нагеля. Она утверждала: многие вопросы о субъективном опыте летучих мышей, которые мы все еще считаем открытыми, зависят от вопросов о нейробиологических деталях, которые пока остаются без ответа. Одним из примеров таковых является функция различных профилей корковой активности у совершающей действия летучей мыши.
Широко известно также утверждение Дэвида Чалмерса (Chalmers 1996), что любой возможный подход к сознанию при помощи описания мозговых процессов оставляет открытым «объяснительный разрыв» между мозговым процессом и свойствами сознательного опыта. ‹14›Это связано с тем, что ни одна теория мозговых процессов не может ответить на «сложный» вопрос: почему именно этот мозговой процесс должен вызывать конкретно этот сознательный опыт? Мы всегда можем вообразить («помыслить / conceive of») вселенную, населенную существами, имеющими эти мозговые процессы, но полностью лишенными сознательного опыта. Теории сознания требуется объяснить, как и почему некоторый мозговой процесс вызывает сознательный опыт, изобилующий всеми теми особенностями, которые мы испытываем. Тот факт, что трудный вопрос остается без ответа, показывает, что мы, вероятно, никогда не получим полного объяснения сознания на уровне нейронных механизмов. Пол и Патрисия Чёрчленды (Paul and Patricia Churchland 1997) предложили следующий вариант обнаружения ошибок и ответ. По их мнению, Чалмерс предлагает концептуальный аргумент, основанный на нашей способности воображать существ, обладающих подобным нашему активным мозгом, но полностью лишенных сознательного опыта. Однако чем больше человек узнает о том, как мозг производит сознательный опыт – и такая литература появляется (некоторые ранние разработки представлены в Gazzaniga 1995) – тем труднее становится представить вселенную, населённую существами с подобными нашим мозговыми процессами, но лишенными сознания. Это – не просто утверждение: Черчленды обращаются к некоторым нейробиологическим деталям. Например, Пол Чёрчленд (Churchland 1995) разрабатывает нейронаучный подход к сознанию, основанный на повторяющихся связях между ядрами таламуса, особенно между «диффузно проекционными» ядрами, такими как внутриламинарные ядра и кора.‹15›
Чёрчленд утверждает, что таламокортикальная рекуррентность объясняет избирательные особенности сознания, влияние кратковременной памяти на сознательный опыт, яркие сновидения во время быстрого сна (фаза БДГ, быстрое движение глаз) и другие «ключевые» особенности сознательного опыта. Иными словами, Черчленды утверждают: когда человек узнает о паттернах активности в этих повторяющихся циклах, он больше не может «вообразить» или «помыслить» эту активность, протекающую без возникновения основных особенностей сознательного опыта. (Помимо простого проговаривания фразы «Я сейчас воображаю деятельность в этих цепях без избирательного внимания / эффектов кратковременной памяти / ярких сновидений /…»).
Второе направление скептических рассуждений о полном нейронаучном объяснении сознания касается сенсорных квалиа: интроспективных качественных аспектов чувственного опыта, – особенностей, благодаря которым субъекты распознают сходства и различия в своем опыте. Цвета в случае зрительных ощущений – любимый пример философов. Одна известная философская головоломка, касающаяся цветовых квалиа – это предположение о мыслимости спектральных инверсий. Многие философы утверждают: концептуально возможно (если, вероятно, даже невозможно физически), чтобы два человека не отличались нейрофизиологически, в то время как цвет пожарных машин и помидоров, видимый одним из субъектов, был бы цветом, который другой субъект распознаёт у травы и лягушек (и наоборот). Большое количество ориентирующихся на нейронауки философских работ обращалось к данному вопросу. (C.L. Hardin 1988 и Austen Clark 1993 – показательные тому примеры.) Смежная с источником возникновения нейрофилософских соображений область касается метафизики самих цветов (нежели цветовых переживаний). Многолетний философский спор заключается в том, являются ли цвета объективными свойствами, существующими вовне тех, кто их воспринимает, или, скорее, сами опознаваемы посредством – или зависимы от – умов или нервных систем воспринимающих субъектов. Определённая нейронаучная работа по этой проблеме начинается с характеристик цветового опыта: например, что суждения о сходстве цветов приводят к упорядочению цветов по кругу (Clark 1993). С опорой на это знание можно приступить к отображению феноменологии на экологические или физиологические закономерности. Если мы отождествим цвета с определенными частотами электромагнитного излучения, структура круга цветовых оттенков не сохранится, в то время как при отождествлении цветов с активностью в нейронах оппонетной обработки – сохранится. Эта любопытная деталь не является решающей для объективистско-субъективистских споров о цвете, но она выражает тип нейрофилософской работы, которая затрагивает традиционные метафизические вопросы, выходящие за рамки философии сознания. (Подробнее описанные проблемы см. в Статье о цвете в SEP.)
Двумя разделами выше в обсуждении работы Хардкасл (Hardcastle 1997) мы увидели, что нейрофилософы вступили на территорию споров о природе и методологическом значении болевых ощущений. Двадцать лет назад Дэниэль Деннетт (Dennett 1978) поднял вопрос о том, возможно ли создать чувствующий боль компьютер. Он сравнивает нейрофизиологические открытия о переживаниях боли и то, что о них говорит здравый смысл, и отмечает напряженность между этими позициями. Он подозревает, что несоизмеримость между научным знанием и здравым смыслом обусловлена непоследовательностью последнего. Установка Деннета – выжидание. Но, предвосхищая ответ Чёрчленда Чалмерсу, Деннетт отдаёт предпочтение научным исследованиям, нежели философским аргументам, основанным на мыслимости (conceivability-based).
Неврологические расстройства привлекли внимание философов, интересующихся сознанием. В течение почти что пятидесяти лет философы обсуждали, какое значение для проблемы единства самости (личности) имеют удостоенные Нобелевской премии эксперименты с пациентами с комиссуротомией, у которых по клиническим причинам было хирургически удалено мозолистое тело‹16› (Nagel 1971). Мозолистое тело представляет собой огромный пучок аксонов, соединяющих нейроны левого и правого полушарий головного мозга у млекопитающих. В тщательно контролируемых экспериментах пациенты с комиссуротомией, судя по всему, демонстрируют два разобщающихся «очага» сознания. Элизабет Шехтер (Schechter 2018) недавно опубликовала значительно обновлённую философскую трактовку научных деталей этих пациентов с «расщепленным мозгом», включив их собственные отчеты о переживаемом опыте, а также проследила, какие последствия для нашего понимания себя влечёт анализ подобных случаев.
В главе 5 своей книги (Churchland 1986) Патриция Черчленд расширила как диапазон, так и философские импликации неврологических расстройств. Одно из расстройств, которые она обсуждает подробно, – это слепозрение. Некоторые пациенты с повреждениями первичной зрительной коры сообщают, что не могут видеть предметы в областях своих зрительных полей, но в испытаниях по вынужденному угадыванию стимулов в этих областях демонстрируют результаты, намного превосходящие шанс случайного угадывания. Для объяснения этого феномена были предложены различные научные и философские интерпретации. Нед Блок (Block 1995) обеспокоен тем, что многие из этих интерпретаций смешивают воедино различные представления о сознании. Он называет эти представления «феноменальное сознание» («P-сознание») и «сознание доступа» («A-сознание»).
Первое – это «каково быть»-ийный аспект сознательных переживаний.
Последнее – это доступность репрезентативного содержания для самостоятельных действий и речи. Блок утверждал, что P-сознание не всегда репрезентативно, в то время как A-сознание – всегда. Деннетт (Dennett 1991, 1995) и Тай (Tye 1993) скептически относятся к нерепрезентативному анализу сознания в целом. Они предоставляют отчеты о слепозрении, не зависящие от введённых Блоком различий.
На этом пункте мы прерываем наш краткий обзор нейрофилософской работы относительно сознания. Многие другие темы также заслуживают нейрофилософского исследования. Мы упомянули комиссуротомию и единство сознания и самости, – темы, которые продолжают вызывать дискуссии. Другие квалиа, помимо переживаний цвета и боли, также быстро привлекли нейрофилософское внимание (Akins 1993a, b, 1996; Austen Clark 1993), как и тема самосознания (Bermúdez 1998).
Локализация когнитивных функций: от изучения поражений до функциональной нейровизуализации
Одна из первых проблем, возникших в неврологии еще в XIX веке, касалась локализации определенных когнитивных функций в определенных областях мозга. Хотя «локализационный» подход имел сомнительное происхождение во френологии Галля и Шпургейма и активно оспаривался Флурансом в начале XIX века, он вновь возник в конце XIX века при исследованиях афазии, которые проводили Буйо, Аубуртин, Брока и Вернике.
Эти неврологи проводили тщательные исследования (когда представлялось возможным) речевых нарушений у пациентов с афазией, после чего посмертно осуществляли вскрытия головного мозга‹17›.
Первоначальное исследование Брока в середине XIX века с участием 22 пациентов подтвердило, что преобладающим было повреждение левого полушария коры, и что для расстройства в производстве речи необходимо повреждение второй и третьей лобных извилин. Хотя предположенные Брока анатомические координаты «центра производства речи» не вполне точно соотносятся с повреждениями, вызывающими расстройства речепорождения, но и эта область лобной коры, и расстройства производства речи по-прежнему носят его имя («область Брока» и «афазия Брока»). Менее чем через два десятилетия Карл Вернике опубликовал свидетельства о существовании второго речевого центра. Эта область анатомически отличается от области Брока, и ее повреждение вызывало совсем другой набор афазических симптомов.
Область коры, которая до сих пор носит его имя («область Вернике»), расположена вокруг первой и второй извилин в височной коре, а названная его именем афазия («афазия Вернике») связана с расстройством понимания языка. Метод Вернике, как и метод Брока, был основан на исследованиях поражений, вызванных естественными травмами: тщательная оценка поведенческих расстройств с последующим посмертным вскрытием для выявления мест повреждения тканей и атрофии. Позднейшие и более тщательные исследования поражений предлагают уточнённую локализацию определенных лингвистических функций, а также и по сей день остаются краеугольным камнем в изучении афазии.
Исследования поражений также позволили получить данные о локализации других когнитивных функций: например, сенсорной обработки и определенных типов обучения и памяти. Впрочем, аргументы о локализации упоминаемых здесь других функций неизменно включают в себя исследования, использующие животные модели. На животной модели можно проводить тщательные поведенческие измерения в строго контролируемых условиях, а затем удалять определенные участки нервной ткани (или использовать множество других методов, чтобы блокировать или усиливать активность в этих областях) и повторно измерять успешность выполнения тех же поведенческих тестов.
Поскольку у нас нет общепринятых животных моделей того, как происходит производство человеческого языка или его понимание, эти дополнительные свидетельства недоступны для неврологов или нейролингвистов. Данное ограничение делает неврологическое изучение языка парадигмальным случаем, позволяющим оценить логику метода повреждения / расстройства как средства определения функциональной локализации. Барбара фон Эккардт (Von Eckardt Klein 1978) попыталась четко обозначить шаги рассуждения, связанные с этим распространенным и исторически важным методом.
Ее анализ начинается с широко известного анализа функционального объяснения, предложенного Робертом Камминсом, но Барбара расширяет его до понятия структурно адекватного функционального анализа. Такого рода анализы разбивают сложную способность C на составляющие ее способности c1 , c2 ,…, cn , где составляющие способности соответствуют базовым структурным деталям системы. Например, производство человеческой речи (сложная способность C) является результатом формулирования речевого намерения (speech intention), затем – выбора соответствующих лингвистических репрезентаций, ухватывающих содержание речевого намерения, затем – формулирования моторных команд для получения соответствующих звуков, а затем – направления этих моторных команд в соответствующие моторные пути (перечисленные выше, все вместе – это составляющие способности c1 , c2 , ..., cn ). Гипотеза о функциональной локализации имеет вид: структура мозга S в организме (определённого типа) O имеет составляющую способность ci , где ci является функцией некоторой части O. Примером может служить утверждение: область Брока (S) у человека (O) формулирует моторные команды для получения соответствующих звуков (одна из составляющих способностей ci ). Подобные гипотезы определяют аспекты того, как структурно реализована функционально-компонентная модель. Они являются частью теории нейронной реализации функциональной модели.
Вооружившись этими формализациями, фон Эккард Кляйн утверждает, что вывод к гипотезе о функциональной локализации происходит в два этапа. Во-первых, формулируется гипотеза о функциональном расстройстве у пациента – на основании аномального поведения, которое пациент проявляет. Во-вторых, локализация функции в нормальном мозге выводится на основе гипотезы о функциональном расстройстве плюс данных о месте повреждения мозга. Структурно-адекватный функциональный анализ способности связывает патологическое поведение с предполагаемым функциональным расстройством. Эта связь предполагает четыре условия адекватности, накладываемые на гипотезу функционального расстройства. Во-первых, патологическое поведение P (например, речевое расстройство, характерное для афазии Брока) должно быть результатом неспособности использовать некоторую сложную способность C (производство человеческой речи). Во-вторых, должен наличествовать структурно-адекватный функциональный анализ того, как люди используют способность C, включающую в себя некоторую составляющую способность ci (формулирование моторных команд для получения соответствующих звуков). В-третьих, действие описанных структурно-адекватным функциональным анализом этапов минус действие компонента, выполняющего ci (область Брока), должно привести к патологическому поведению P. В-четвертых, не должно наличествовать лучшего доступного объяснения того, почему пациент делает Р.
Аргументы в пользу гипотезы функционального расстройства на основе патологического поведения, таким образом, являются примером аргументации в пользу наилучшего доступного объяснения. Когда постулирование расстройства в нормальном функциональном компоненте обеспечивает наилучшее доступное объяснение патологических данных, совершаемый нами вывод оказывается оправдан.
Фон Эккардт Кляйн применяет этот анализ к неврологическому исследованию, включающему противоречивую переинтерпретацию агнозии‹18›. Ее философское объяснение этого важного неврологического метода показывает, что большинство вызовов аргументам локализации либо спорят только против локализации определенного типа функциональной способности, либо против обобщения от локализации функции у одного человека до всех нормальных людей. (Она приводит примеры и того, и другого из неврологической литературы.)
Такие проблемы не ставят под сомнение обоснованность стандартных аргументов в пользу функциональной локализации на основании повреждений. Из этого не следует, что такие аргументы беспроблемны.
Но они сталкиваются со сложными фактологическими и методологическими проблемами, а не логическими. Кроме того, анализ этих аргументов, включающий тип функционального анализа и вывод к наилучшему из доступных объяснений, имеет важное значение для биологического изучения когнитивной функции. Функциональный анализ требует функциональных теорий, а структурно адекватный функциональный анализ требует проверок, налагаемых науками более низкого уровня, исследующими основные физические механизмы. Аргументы в пользу лучшего доступного объяснения часто затрудняются отсутствием теоретического воображения: доступные альтернативные объяснения часто сильно ограничены. Мы должны искать теоретическое вдохновение на любом уровне исследования или объяснения.
Следовательно, делая явным «логику» этой распространенной и исторически важной формы неврологического объяснения, выявляется необходимость совместного участия на всех научных уровнях, от когнитивной психологии до молекулярной нейронауки. Таким образом, фон Эккардт Кляйн (Von Eckardt Klein 1978) предвосхитила то, что стало называться «методологией коэволюционных исследований», которая остается центральной частью нейрофилософии до наших дней (см. Раздел 6).
За последние три десятилетия новые данные о локализации когнитивных функций все чаще поступают из нового источника: разработок и совершенствования методов нейровизуализации. Тем не менее, логическая форма аргументов о локализации функций, по-видимому, не изменилась по сравнению с теми, которые используются в исследованиях поражений и которые проанализировала фон Эккард Кляйн. Различие в том, что эти новые технологии нейровизуализации решают некоторые методологические проблемы, с которыми сталкивались исследования поражений. Например, исследователям больше не нужно ждать, пока пациент умрет, при этом рискуя дополнительно повредить головной мозг в поисках исходного места повреждения.
В философских дискуссиях были выделены два многообещающих метода функциональной визуализации: позитронно-эмиссионная томография (ПЭТ) и функциональная магнитно-резонансная томография (фМРТ). Несмотря на то, что они измеряют различные биологические маркеры функциональной активности, одобренная для использования на людях ПЭТ теперь имеет пространственное разрешение вплоть до диапазона в 1 мм, в то время как фМРТ имеет разрешение вплоть до расстояний менее 1 мм.‹19›
Поскольку эти методы увеличивали пространственное и временное разрешение функциональных маркеров и продолжали использоваться вместе со сложными поведенческими методологиями, аргументы в пользу локализации специфических психологических функций во все более конкретизируемых нервных областях продолжали культивироваться. Стаффльбим и Бехтел были одними из первых, кто предоставил философски содержательное обсуждение ПЭТ. Бехтел и Ричардсон (Bechtel and Richardson 1993) предоставили общую основу для аргументов «локализации и декомпозиции», которая во многом предвосхитила «новую механистическую» перспективу в философии науки и философии нейронаук (см. Разделы 7 и 8 далее). Бехтел и Мандейл (Bechtel and Mundale 1999) провели дальнейшее уточнение адаптированных для нейронаук философских аргументов в пользу локализации функций.
Сравнительно недавнее философское обсуждение описанных функциональных методов визуализации было склонно призвать к большей осторожности в том, чтобы утверждения о локализации опирались на результаты [этих методов визуализации]. К примеру, Роскис (Roskies 2007) указывает на склонность думать о доказательной силе функциональных нейроизображений (особенно фМРТ) по аналогии с фотографиями. Опираясь на работу в области эстетики и изобразительного искусства, Роскис утверждает: многие из функций, которые придают фотографиям их доказательную силу, отсутствуют в функциональных нейроизображениях.
Таким образом, хотя нейроизображения и служат доказательством для заявлений о нейрофункциях и даже для гипотез локализации, нюансы их правильной интерпретации гораздо сложнее, чем подчас предполагают философы. Что ещё важнее, Клейн (Klein 2010) утверждает: возникающие в результате функциональной нейровизуализации (особенно фМРТ) изображения «мозговой активности» являются плохим доказательством функциональных гипотез.
Для этих изображений обычно представлены результаты тестирования на данных фМРТ значимости нулевой гипотезы, но одно лишь такое тестирование не может предоставить доказательства функциональной структуры в случае такой причинно-плотной системы, какой является человеческий мозг. Вместо этого правильный способ интерпретации функциональных нейроизображений – указание на области, относительно которых оправданы дальнейшие данные и анализ. Однако эти данные, как правило, требуют отнюдь не только простого тестирования значимости, поэтому скептицизм в отношении доказательной силы нейроизображений не оправдывает скептицизм в отношении фМРТ в целом.
Локализация функций и по сей день остается центральной дискуссионной темой в философии нейронаук. Мы рассмотрим более актуальные работы в нижеследующих разделах.
Результат коэволюционной исследовательской идеологии: фокусировка философии на когнитивной и вычислительной нейробиологии
Сегодняшние нейронаучные открытия, касающиеся клеточных и молекулярных механизмов нейронной проводимости и передачи сигналов, поистине впечатляют. Эти результаты воплощают собой одно из высочайших достижений научного поиска. (Сомневающимся советуем просто в течение пяти минут полистать недавно вышедший том «Общества нейронаук: аннотации» / Society for Neuroscience Abstracts.) Менее всеобъемлющими, но не менее впечатляющими являются открытия на «более высоких» уровнях нейронаук: уровнях цепей, сетей и систем.
Все эти достижения являются естественным результатом растущей научной специализации. Чтобы раздвинуть границы своего понимания природы, мы развиваем технологии, экспериментальные методы и, в конечном счете, ориентированные на результаты экспериментальные теории – и всё это в рамках конкретных дисциплин. Тем не менее, зачастую из вида теряют важнейший аспект общей картины: взаимосвязь между уровнями, – «клей», который, с одной стороны, связывает знания о деятельности нейронов с их субклеточными и молекулярными механизмами «внизу», а с другой – со схемами активности цепей, сетей и систем «наверху» ». Эта проблема особенно остро встаёт в тех случаях, когда мы пытаемся донести «когнитивистские» психологические теории, – постулируя описание в виде информационных представлений и процессов, действующих над их содержимым, – до уровня нейрональной активности.
«Коэволюция» между этими уровнями объяснения все еще кажется скорее отдаленной мечтой, нежели оперативной методологией, которой можно было бы руководствоваться в повседневных научных исследованиях.
Именно здесь некоторые философы и нейроучёные обратились к вычислительным методам (Churchland and Sejnowski 1992). Одна из надежд заключалась в том, что способ, которым вычислительные модели функционируют в более развитых науках, – вроде физики, – сам сможет оказаться полезной моделью работы. Одним из вычислительных ресурсов, который успешно применяется в более развитых науках для решения аналогичных «межуровневых» задач, являются динамические системы. К глобальным явлениям – вроде крупномасштабных метеорологических паттернов – оказалось продуктивным обратиться как к динамическим, нелинейным и зачастую хаотическим взаимодействиям между физическими явлениями более низкого уровня. Рассмотрение взаимосвязанных уровней теории и объяснения в случае сознания / мозга с использованием вычислительных ресурсов, которые сработали для объединения уровней в более зрелых науках, может дать сопоставимые результаты. Эта методология непременно является междисциплинарной и опирается на ресурсы и исследователей различных уровней, включая высшие – такие, как экспериментальная психология, искусственный интеллект и философия науки.
Использование вычислительных методов в самой нейробиологии не ново. В гигантском аксоне кальмара Ходжкин, Хаксли и Кац (Hodgkin, Huxley, Katz 1952) экспериментально измерили значения зависящей от напряжения проводимости натрия и калия, и включили их в уравнение из физики, описывающее временную эволюцию кинетического процесса первого порядка. Это уравнение позволило им рассчитать наиболее подходящие кривые для смоделированных данных, описывающих проводимость как функцию от времени, а также воспроизвести изменение мембранного потенциала спустя некоторое время после того, когда генерировались потенциалы действия. Также, используя заимствованные из физики уравнения, Ролл (Rall 1959) разработал кабельную модель дендритов. Эта модель обеспечила подход, согласно которому различные входные данные по длине дендритного дерева взаимодействуют во времени и в пространстве, определяя свойства ввода-вывода отдельных нейронов. Данная модель остается влиятельной и сегодня – она была включена в программное обеспечение GENESIS, на котором программируются реалистичные нейроноподобные сети (Bower and Beeman 1995; см. обсуждение в Разделе 2 выше). Дэвид Спаркс и его коллеги показали, что построенная на усреднении векторов модель активности нейронов верхних колликулов правильно предсказывает экспериментальные результаты относительно амплитуды и направления саккадических движений глаз (Lee, Rohrer, Sparks 1988). Работая с более сложной математической моделью, Апостолос Георгопулос и его коллеги предсказали направление и амплитуду движений рук и кистей на основе усредненной активности 224 клеток в моторной коре. Их прогнозы были подтверждены различными экспериментальными испытаниями (Georgopoulos, Schwartz, Kettner 1986). Мы упоминаем эти конкретные исследования лишь потому, что знакомы с ними. Без сомнения, мы легко могли бы увеличить число примеров плодотворного взаимодействия вычислительных и экспериментальных методов в нейробиологии во сто крат. Многие из них сформировались еще до того, как «вычислительная нейробиология» стала признанной областью приложения исследовательских усилий.
Мы уже видели один пример, – подход преобразования векторов к описанию нейронных представлений и вычислений, – когда-то активно развивавшийся в когнитивной нейробиологии (см. Раздел 2 выше). Другие подходы, использующие «когнитивистские» ресурсы, были и продолжают применяться‹20›.
Некоторые из этих проектов опираются на «когнитивистские» характеристики подлежащих объяснению явлений. Некоторые используют «когнитивистские» экспериментальные техники и методологии. Некоторые даже пытаются вывести «когнитивистские» объяснения из клеточно-биологических процессов (например, Hawkins and Kandel 1984). Как выразился Стивен Косслин (Kosslyn 1997), когнитивные нейробиологи смотрят на сознание из характерно-когнитивистской перспективы «обработки информации», не пытаясь отделить её от теорий о механизмах работы мозга. Подобное начинание нуждается в том, чтобы междисциплинарное сообщество и само стремилось бы передать заинтересованным неспециалистам релевантные данные из того колоссального их объёма, который собран в отдельных дисциплинах. Для этого требуется нечто большее, чем одни люди, желающие общаться с другими, работающими на смежных уровнях: также нужны исследователи, специально обученные методам и знанию фактических деталей из различных дисциплин. Подобная потребность обескураживает, но она дает надежду философам, желающим внести свой вклад в реальные нейронауки. Мыслители, обученные как даруемому философией «обзорному видению», так и научной и экспериментальной основам подлинной науки (уровня выпускника), были бы идеально оснащены для решения этой задачи. До расцвета программам магистратуры по философии саму эту потенциальную нишу распознавали сравнительно медленно, однако некоторые образовательные программы предприняли шаги по её занятию (см., например, раздел Другие интернет-ресурсы в SEP).
Впрочем, остается один явный недостаток. Учитывая подготовку и интересы философов, именно «высокоуровневые» нейронауки – цепи, когнитивные системы и объединяющиеся с ними области вычислительной нейробиологии – склонны больше всего привлекать философское внимание. Сколь естественным не был бы подобный фокус, он может привести философов к ошибочной картине нейронаук. Нейробиология остается сфокусированной на клеточных и молекулярных механизмах нейрональной активности и сотрудничает с поведенческой нейробиологией, работающей с животными моделями.
Именно таким образом большинство членов Общества нейронаук, в настоящее время включающем свыше 37 000 членов, по-прежнему классифицирует свои собственные исследования; именно туда направляется большая часть денег, идущих на грантовые исследования; и, наконец, экспериментальные публикации именно в этих областях чаще всего появляются в наиболее цитируемых научных журналах. (ссылка на веб-сайт Общества нейронаук / the Society for Neuroscience - в разделе Другие интернет-ресурсы ведёт к большому массиву данных о соответствующей статистике; см., в частности, раздел «Публикации».)
И все же философы не уделяют большого внимания клеточной и молекулярной нейробиологии.
К счастью, сейчас эта ситуация, по-видимому, меняется, о чем мы расскажем в нижеследующих Разделах 7 и 8. Тем не менее, преобладающее внимание, уделяемое философами когнитивной / системной / вычислительной нейробиологии, затеняет важность экспериментально-ориентированного лабораторного фокуса исследований в современной нейробиологии.
Достижения в философии нейронауки
Различие между «философией нейронаук» и «нейрофилософией» начало становится отчётливее в течение первого десятилетия XXI века – в первую очередь, благодаря большему количеству вопросов, рассматриваемых в обеих областях. В философии нейронаук по-прежнему есть тенденция ставить традиционные вопросы из области философии науки, применяя их конкретно к нейронаукам. К таким вопросам относятся: какова природа нейронаучного объяснения? А также – какова природа открытия в нейронауках?
Ответы на эти вопросы рассматриваются либо в описательном ключе (как функционирует нейронаука?), либо в нормативном (как должна развиваться нейронаука)? Некоторые нормативные проекты в философии нейронаук являются «деконструктивными»: они критикуют выдвигаемые нейроучёными заявления по различным темам. Например, философы нейронаук подвергли критике концепцию личности, которая была принята у исследователей в когнитивной нейробиологии (см. Roskies 2009). Другие нормативные проекты носят конструктивный характер, предлагая новые теории нейрональных явлений или методы интерпретации нейронаучных данных.
Такие проекты часто плавно интегрируются с самой теоретической нейронаукой. Например, Крис Элиасмит и Чарльз Андерсон разработали подход к построению нейрокомпьютерных моделей в своей книге «Нейронная инженерия» (Eliasmith and Anderson 2003). В отдельных публикациях Элиасмит утверждал, что введенная в «Нейронной инженерии» базовая схема предоставляет как нормативный отчет о нейронных репрезентациях, так и схематизм для унификации объяснений в нейронауках (например, Eliasmith 2009).
Нейрофилософия же продолжала применять находки из областей нейронаук к традиционным философским вопросам. Тому есть множество примеров: что такое эмоция? (Prinz 2004) Какова природа желания? (Schroeder 2004) Как социальное познание стало возможным? (Goldman 2006) Что является нейронной основой морального познания? (Prinz 2007) Что является нейронной основой счастья? (Flanagan 2009) Нейрофилософские ответы на эти вопросы ограничены сведениями, которыми нейронауки располагают относительно нервной системы. Например, в своей книге « Три лика желания» Тимоти Шрёдер (Schroeder 2004) утверждал, что наша общепринятая концепция желания приписывает данному состоянию три способности:
- (1) способность подкреплять поведение при удовлетворении,
- (2) способность мотивировать поведение и
- (3) способность определять источники удовольствия. Основываясь на публикациях о функции дофамина и теории обучения с подкреплением, Шрёдер утверждал, что обработка вознаграждений является основой для всех трех перечисленных способностей.
Таким образом, сущность желания - это вознаграждение.
В течение первого десятилетия XXI века в нейрофилософии возникла тенденция искать в нейронауках руководство в сфере моральной философии. Это должно быть очевидно по только что упомянутым нами темам. В то же самое время возобновился и интерес к морализации относительно нейронаук и неврологических методов лечения (см. Levy 2007; Roskies 2009). Соответственно, эта новая область, нейроэтика, объединила в себе как интерес в релевантности данных нейронаук для понимания морального познания, так и релевантность моральной философии для получения и регулирования применения знаний из нейронаук. Регуляторная ветвь нейроэтики изначально была явно сосредоточена на этике лечения людей, страдающих неврологическими нарушениями, этике попыток улучшить когнитивные способности человека (Schneider 2009), этике применения технологии «чтения мыслей» к проблемам в криминалистике (Farah and Wolpe 2004), а также этике экспериментов на животных в нейронауках (Farah 2008). В последнее время обе эти области нейроэтики переживают период колоссального роста. Заинтересованный читатель должен обратиться к статье по нейроэтике в наших переводах «SEP на русском».
Наблюдавшиеся в течение первого десятилетия XXI века в философии нейронаук тенденции включали в себя возобновление интереса к природе механистических объяснений. Это соответствовало общему тренду в философии науки (например, см. Machamer, Darden и Craver 2000). Применение этого общего подхода и к нейронаукам неудивительно. Термин «механизм» широко используется среди нейроучёных. В своей книге «Объясняя мозг» (Craver 2007) Карл Крейвер утверждал, что механистические объяснения в нейробиологии являются причинными объяснениями, причём, как правило, многоуровневыми. Например, объяснение нейронального потенциала действия включает в себя: сам потенциал действия; клетку, в которой он возникает; электрохимические градиенты и, наконец, белки, сквозь которые ионы проходят через мембрану. Таким образом, мы имеем сложноставную сущность (клетку), причинно взаимодействующую с нейротрансмиттерами на ее рецепторах. Компоненты клетки участвуют в различных действиях, - например, в открытии и закрытии лиганд-управляемых и управляемых напряжением ионных каналов, чтобы создать паттерн изменений: деполяризующий ток, составляющий сам потенциал действия. Таким образом, механистическое объяснение потенциала действия охватывает сущности на клеточном, молекулярном и атомном уровнях, каждые из которых имеют причинное отношение к созданию потенциала действия. Описанная причинно-следственная связь может быть подтверждена посредством изменения любой из этих переменных, - например, плотности ионных каналов в клеточной мембране, чтобы сгенерировать изменение в потенциале действия; подтверждение причинно-следственной связи также возможно за счёт проверки согласованности предполагаемой инвариантности между переменными. Вызовы, стоящие перед Крейверовским подходом к механистическому объяснению в нейронауках, особенно в отношении потенциала действия, см. в работах Weber 2008 и Bogen 2005.
Согласно эпистемическим нормам, неявно разделяемым нейроучёными, хорошие нейронаучные объяснения – это хорошие механистические объяснения; в свою очередь, хорошими механистическими объяснениями считаются такие, которые ухватывают инвариантные отношения между механизмами и контролируемыми ими явлениями. (Более полное описание инвариантности в причинных объяснениях в науке см. в работе Джеймса Вудворда : Woodward 2003. Механицисты широко используют «интервенционистский» подход Вудворда к причинам и причинным объяснениям.) Предложенный Крейвером подход поднял вопросы о месте редукции в нейронауках. Джон Бикл (Bickle 2003) предположил, что рабочая концепция редукции в нейронауках заключается в обнаружении систематических взаимосвязей между вмешательствами на более низких уровнях биологической организации, – как это делается в клеточной и молекулярной нейробиологии, – и на более высоких уровнях поведенческих следствий, – как это описывается в психологии. Бикл назвал данную перспективу «практическим редукционизмом (reductionism-in-practice)», чтобы обозначить её отличие от концепций интертеоретической или метафизической редукции, которые были в центре многих дискуссий в философии науки и философии сознания.
Впрочем, несмотря на переформулировку Биклом редукции, механицисты обычно сопротивляются или, по крайней мере, релятивизируют ярлык «редукционизма». Крейвер (Craver 2007) называет свою точку зрения «мозаичным единством» нейронаук. Бехтел (Bechtel 2009) называет свою позицию «механистической редукцией(-измом)». И Крейвер, и Бехтел защищают перспективу многоуровневых «механизмов-внутри-механизмов», в которой нет эпистемически привилегированного уровня механизмов. Это отличает их от редукции(-онизма), безжалостно или на иных основаниях отдающего преимущество более низким уровням. Тем не менее, мы можем спросить: является ли механизм своего рода практическим редукционизмом? Или механизм, рассмотренный как позиция в нейронаучном объяснении, всё же предполагает некоторую автономию психологии? Если он предполагает автономию, редукционисты могут бросить вызов механицистам по поводу подобного предположения. С другой стороны, практический редукционизм Бикла явно отходит от интертеоретической редукции в смысле, в котором последняя понимается в философии науки. Как признает сам Бикл, последний его проект редукционизма был в значительной степени вдохновлен критикой механицистов в адрес его предыдущего подхода «новой волны». Механицисты могут предъявить Биклу претензию, что его отдаление от традиционных подходов также привело к отдалению от интересов, которые мотивировали сами эти подходы. (См. Polger 2004 касательно соответствующих претензий.) Как мы увидим в нижеследующем Разделе 8, эти проблемы, связанные с механистической философией нейронаук, стали более актуальными, поскольку механизм стал доминирующим представлением в этой области.
Роль временной репрезентации в сознательном опыте и виды нейронных архитектур, достаточные для представления объектов во времени, также вызвали интерес. В традиции феноменологии Гуссерля, Дэн Ллойд (Lloyd 2002, 2003) и Рик Граш (Grush 2001, 2009) порознь обратили внимание на трехчастную временную структуру феноменального сознания как того, что подлежит нейронаучному объяснению. Эта структура состоит из субъективного настоящего, непосредственного прошлого и ожидания непосредственного будущего. Например, сознательное осознавание мелодии относится не просто ко временному срезу впечатлений от мелодии, но к ноте, присутствовавшей минуту назад, другой, которая присутствует сейчас, и ожиданию последующих нот в непосредственном будущем. По мере того, как этот опыт продолжается, то, что мгновение назад было непосредственным во времени, теперь сохраняется как мгновение ближайшего прошлого; то, что ожидалось (произойдёт нечто или нет), теперь стало переживаемым настоящим; и также сформировалось новое ожидание о том, что будет дальше. Опыт человека не является статичным, даже если это опыт лишь одного (мелодии). Упомянутые ранние работы обрели большую актуальность с появлением моделей, описывающих функционирование всего мозга на основе «предиктивного кодирования». Менее чем за десять лет после публикаций Ллойда и Граша модели «предиктивного кодирования» были разработаны нейроучёными, в числе которых был Карл Фристон (Friston 2009), а также получили более широкое философское освещение благодаря Якобу Хоуи (Hohwy 2013) и Энди Кларку (Clark 2016).
По словам Ллойда, трехчастная структура сознания ставит уникальную проблему перед анализом данных фМРТ и планированием экспериментов. Проблема возникает из-за напряженности между тождеством объекта опыта (например, развитие той же самой мелодии) и временной подвижностью самого опыта (например, переходов между услышанными нотами). В то время, когда Ллойд писал свою работу, одно из стандартных средств анализа данных фМРТ заключалось в усреднении нескольких наборов данных и вычитании предполагаемого значения базовой активации из общего состава.
Это делалось, чтобы отфильтровать шумы от связанного с задачей гемодинамического отклика. Однако, как указывает Ллойд, эта распространенная в то время практика игнорирует большую часть данных, необходимых для изучения нейронных коррелятов сознания.
Она создает статические изображения, которые пренебрегают отношениями между измеряемыми параметрами с течением времени. Вместо этого Ллойд применяет многовариантный подход к изучению данных фМРТ, предполагая, что в основе временной обработки, порождающей переживаемое время, лежит рекуррентная сетевая архитектура. Простая рекуррентная сеть имеет входной слой, выходной слой, скрытый слой и дополнительный слой, который копирует предыдущее состояние активации либо скрытого слоя, либо выходного слоя. Позволяя выходному слою представлять прогнозируемый результат, входной слой может затем представлять текущее состояние, а дополнительный уровень – предыдущее состояние. Такое назначение имитирует трехчастную структуру опыта времени посредством сетевой архитектуры.
Если лежащие в основе сознательного опыта нейрональные механизмы можно приблизительно выразить рекуррентной сетевой архитектурой, одно из предсказаний состоит в том, что текущие состояния нейронов несут информацию о ближайших будущих и предыдущих состояниях.
Применительно к фМРТ модель предсказывает, что временные точки в серии изображений будут содержать информацию о предыдущих и последующих временных точках. Результаты проведённого Ллойдом (Lloyd 2002) анализа наборов данных о 21 субъекте, взятых из общедоступного Национального центра данных фМРТ, подтверждают это предсказание.
Интерес Граша (Grush 2001, 2004) к временным репрезентациям является частью его более обширного, систематического проекта по решению семантической проблемы в вычислительной нейробиологии, а именно: как мы можем отграничить изучение мозга как информационного процессора от изучения любого другого сложного причинного процесса? Этот вопрос возвращает нас к знакомой территории психосемантики (см. Раздел 3 выше), но теперь отправная точка располагается внутри практик вычислительной нейробиологии.
Таким образом, семантическая проблема становится задачей для философии нейронаук, поскольку она вопрошает нас: что означает (или должно означать) «вычисление» в вычислительной нейробиологии?
Решение Граша основывалось на концепциях из современной теории управления. В дополнение к контроллеру, датчику (сенсору) и целевому состоянию в некоторых типах систем управления задействуется модель процесса, которая моделирует текущий контролируемый процесс. Модель процесса способна облегчать осуществление различных инженерных функций, включая преодоление задержек в обратной связи и фильтрацию шумов. Точность модели процесса можно оценить относительно ее «совместимости по соединителям (plug-compatibility)» с реальным процессом. Совместимостью по соединителям считается мера того, в какой степени контроллер способен причинно связаться с моделью процесса для получения тех же результатов, которые он мог бы получить, соединившись с реальным процессом. Обратите внимание, что совместимость по соединителям не является информационным отношением.
Чтобы проиллюстрировать потенциальную нейронаучную реализацию, Граш рассматривает в качестве контроллера часть моторных систем мозга (например, премоторную кору). Датчиками являются органы чувств (например, рецепторы растяжения мышц). Модель процессов костно-мышечной системы может существовать в мозжечке (см Kawato 1999). Если контролирующая часть двигательной системы посылает пакеты импульсов в мозжечок точно так же, как она посылает импульсы в костно-мышечную систему, - и, если, в свою очередь, мозжечок принимает пакеты импульсов, аналогичные реальной периферической обратной связи, - тогда мозжечок эмулирует костно-мышечную систему (до такой степени, что фиктивная обратная связь напоминает реальную периферийную обратную связь). Предлагаемое целое, в пределах которого осуществляются вычислительные операции, - это и есть нейронная реализация модели процесса и ее компонентов, или, в терминах Граша, «эмулятора» и его «артикулянтов».
Детали предлагаемой Грашем схемы слишком сложны, чтобы можно было представить их в коротком пересказе. (Например, потому что он вводит множество концептуальных средств для обсуждения репрезентации внешних объектов.) Но, говоря вкратце, он утверждает: понимание временных репрезентаций начинается с понимания того, как эмулируется синхронизация непредвиденных сенсомоторных событий. Успешное последовательное поведение (например, ловля рыбу с помощью копья) зависит не только от отслеживания того, где человек находится в пространстве, но и от того, где он находится во временном порядке движений, а также временного расстояния между текущими, предыдущими и последующими движениями. Выполнение последующего движения может зависеть от отслеживания того, было ли предыдущее движение успешным и соответствует ли текущее движение предыдущим ожиданиям. Граш основывает свои рассуждения на эмуляторах, – моделях процессов в центральной нервной системе, – которые прогнозируют, сохраняют и обновляют фиктивную сенсомоторную обратную связь, синхронизируя их выходной сигнал пропорционально обратной связи от реального процесса (Grush 2005).
Подходы Ллойда и Граша к изучению временных репрезентаций различаются по своим акцентам. Тем не менее, их объединяет неявная приверженность локализации когнитивных функций и разложению их на подфункции посредством ограничений как «сверху вниз», так и «снизу вверх». (См. работу Bechtel and Richardson 1993, где эта общая объяснительная стратегия описывается более подробно) Как мы упоминали несколькими абзацами выше, оба автора разработали важные и интересные способы рассуждения, в которых предвосхитили более свежие нейронаучные и философские работы, посвящённые предиктивному кодированию и мозгу. Оба они создали механистические объяснения, мало беспокоясь о дисциплинарных границах. Один из главных уроков, который можно вынести из работ Бикла и Крейвера, заключается в том, что подобным образом структурирована и нейронаучная практика в целом. Онтологические последствия принятия этого подхода продолжают обсуждаться.
Достижения за второе десятилетие XXI века
Понятие механизма, рассмотренное нами в Разделе 7 выше, стало доминирующим в философии нейронаук на протяжении всего второго десятилетия XXI века. Одним из широко обсуждаемых примеров его использования является работа Гуалтьеро Пиччинини и Карла Крейвера (Piccinini and Craver 2011).
Авторы используют два популярных механистических понятия. Первое из них – многоуровневая перспектива вложенных иерархий механизмов-внутри-механизмов, восходящая к работе (Craver and Darden 2001) и которую мы обсуждали в Разделе 7 выше. Второе – это понятие «наброска механизма», первоначально предложенное в работе (Machamer, Darden and Craver 2000) и детально разработанное в (Craver 2007). Цель Пиччинини и Крейвера состоит в том, чтобы «бесшовно» разместить психологию в составе «интегрированной схемы» наряду с нейронауками.
Они интерпретируют знакомые в психологии функциональные анализы когнитивных способностей как относительно неполные механизмы-наброски, опускающие многие компоненты других механизмов в своём составе, которые в конечном итоге полностью объясняют поведение системы.
Нейронауки, в свою очередь, заполняют места этих недостающих компонентов, динамических процессов и организационных структур – по крайней мере, теми, которые обнаруживаются в нервной системе. Соответственно, это заполнение превращает механизмы-наброски психологии в полномасштабные механистические объяснения.
Таким образом, хотя психология и исходит из функциональных анализов, последние, будучи проинтерпретированы в предложенной схеме, являются механистическими. Пиччинини и Крейвер понимают, что их «интегрированный» подход вступает в противоречие с классическими притязаниями психологии на «автономию» по отношению к нейронаукам. Как бы то ни было, авторы настаивают: вызов, который они бросают классической «автономии» не обязывает их склониться к «редукционизму» ни в его классическом, ни в более свежем варианте. Их приверженность вложенной иерархии механизмов-внутри-механизмов для учета поведения системы признает важность механизмов и причинно-следственных связей на всех составляющих систему уровнях, а не только на более низких (т.е., клеточных и молекулярных).
Дэвид Каплан и Крейвер (Kaplan and Craver 2011) устремляют критический взгляд из механицистской перспективы на математические модели динамических систем, снискавшие популярность в современных системной и вычислительной нейробиологии. Они утверждают, что такие модели имеют объяснительную силу, только если существует «правдоподобное отображение» между элементами модели и элементами моделируемой системы. По сути, речь идёт о том, что на объяснение накладывается ограничение «отображения модели на механизм» (Model-to-Mechanism-Mapping, 3M). Переменные в реально объяснительной модели соответствуют компонентам, действиям или организационным свойствам объясняемой системы. А, в свою очередь, зависимости между переменными модели, обычно получающие математическое выражение в системной и вычислительной нейробиологии, соответствуют причинно-следственным связям между компонентами системы. Каплан и Крейвер оправдывают ограничение 3М на основе объяснительных норм, общих для науки и здравого смысла. При прочих равных условиях, настаивают они, объяснения, предоставляющие более релевантную информацию о компонентах, действиях и организации системы, вероятнее ответят на больше число вопросов о том, как система будет вести себя в различных обстоятельствах, нежели это сделает объяснение, предоставляющее меньше (механистических) деталей. Прилагательное «релевантную» здесь относится к функционированию конкретного механизма. Таким образом, нарушающие 3М-ограничение модели в системной и вычислительной нейробиологии разумнее рассматривать как математические описания явлений, а не объяснения некоторого «немеханистического» разнообразия.
Каплан и Крейвер подвергают испытанию свою собственную позицию с помощью одной из самых популярных динамических / математических моделей во всей вычислительной нейробиологии: модели Хакена-Келсо-Бунца (Haken-Kelso-Bunz 1985) бимануальной координации движений пальцев человека. Авторы указывают на отрывки из публикаций разработчиков модели, позволяющих понять, что разработчики сами намеревались сделать свою динамически-системную модель всего лишь математически компактным описанием временной эволюции «переменной, зависящей исключительно от поведения». Разработчики модели не интерпретировали ни одну из переменных или параметров модели как отображение на компоненты или операции какого-либо гипотетического механизма, генерировавшего бы поведенческие данные. Не подразумевали они также, чтобы какие-либо присутствующие в модели математические отношения или зависимости между переменными отображались в предположенных причинных взаимодействиях между компонентами или действиями какого-либо механизма. Как далее указывают Каплан и Крейвер, после публикации своей динамической модели, сами её разработчики начали исследовать, как описанные в их модели поведенческие закономерности могут быть обусловлены компонентами, действиями и организацией нервной двигательной системы. Их собственное дальнейшее исследование позволяет предположить, что разработчики рассматривали свою динамическую модель как эвристическую: призванную помочь нейроучёным перейти к механистическим объяснениям того, «как это возможно» и, в конечном итоге, к «как оно на самом деле».
По сути, разработанное Капланом и Крейвером 3M-ограничение на объяснение представляет собой дилемму для сторонников динамического подхода. В той мере, в какой разработчики моделей динамических систем намереваются моделировать гипотетические нейронные механизмы исследуемого явления, их объяснения должны согласовываться с 3М-ограничением (и другими канонами механистического объяснения). В той мере, в какой это не является целью разработчиков динамических моделей, их модели не кажутся подлинно объяснительными, - по крайней мере, в определённом смысле «объяснения», характерном для истории науки. Кроме того, когда модели динамических систем оцениваются как успешные, они часто побуждают к последующему поиску базовых механизмов, - так же, как 3M-ограничение и подход механицизм в целом советуют переходить от механизма «как это возможно» к механизму «как он на самом деле». В противном случае «рога» обозначенной выше дилеммы пронзают сторонников подхода динамических систем, утверждающих, что их модели представляют собой необходимый и дополнительный к механистическому вид объяснения в нейронауках (помимо какой бы то ни было эвристической ценности, которую такие модели могут предложить для открытия механизмов).
Радикальный вывод Каплана и Крейвера о том, что динамические «объяснения» являются подлинными объяснениями только в той степени, в которой соблюдают (механистическое) 3М-ограничение, сам должен быть более обоснован. Бремя доказательства всегда лежит на тех, чьи выводы противоречат распространённым допущениям. Требуется нечто большее, чем обсуждение пары знаковых моделей из нейронауки, выполненных в рамках подхода динамических систем (в 2011 году Каплан и Крейвер также обсуждали модель разности гауссианов ‹21›, описывающую свойства рецептивного поля зрительных нейронов млекопитающих). Как можно было ожидать, приверженцы подхода динамических систем взялись за эту задачу. Например, Майкл Зильберштейн и Энтони Чемеро (Silberstein and Chemero 2013) утверждают, что для механистического объяснения характерны стратегии локализации и декомпозиции – и что некоторые объяснения в системной нейробиологии нарушают одно или оба из этих допущений. Подобные нарушения, в свою очередь, создают дилемму для механицистов. Последние либо должны «расширить» своё понимание объяснения за пределы только лишь декомпозиции и локализации (чтобы включить и эти выбивающиеся случаи), либо должны принять «контрпримеры» в состав механистического объяснения в принципе – как в системной нейробиологии, так и в системной биологии в целом.
Лорен Росс (Ross 2015) и Мазвиита Чиримуута (Chirimuuta 2014) независимо друг от друга обращаются к подходу Роберта Бэттермана о минимальной модели объяснения как важному типу немеханистического объяснения в нейронауках. Первоначально, минимальные модели были разработаны для характеристики типов объяснений в физических науках (см., например, Batterman and Rice 2014). В подходе Бэттермана проводится различие между двумя видами вопросов «почему?» в науке: почему явление проявляется в определенных обстоятельствах; и – почему явление проявляется вообще, или в ряду различных обстоятельств. Механистические объяснения отвечают на первый тип вопроса почему. Здесь действует предположение «чем больше деталей, тем лучше / more details the better, MDB» (Chirimuuta 2014), сродни предположению Каплана и Крейвера «при прочих равных» о лучших объяснениях (которое упоминалось выше). Однако минимальные модели, сводящие к минимуму представленные детали внедрения (implantation details) и, следовательно, нарушающие MDB, способны лучше ответить на второй тип научных вопросов «почему». Цитируя вычислительных нейробиологов Ринзеля и Эрментроута, Росс (Ross 2015) настаивает на том, что модели, содержащие слишком много деталей сверх необходимого, неизбежно затеняют распознавание критических значимых элементов, так как оставляют слишком много открытых возможностей, - особенно когда кто-то пытается ответить на вопросы второго типа по Бэттерману о поведении системы.
И Чиримуута, и Росс обращаются к смежным ресурсам вычислительной нейробиологии, чтобы проиллюстрировать применимость Бэттермановской стратегии минимальной модели объяснения. Росс обращается к «каноническим моделям», которые представляют «общие качественные признаки ряда отдельных нейронных систем» (2015: 39). Её центральным примером является вывод модели Эрментрута-Копелла, описывающей возбудимость нейронов I класса, в которой используются «математические методы абстракции» для «редукции моделей молекулярно различных нейронных систем к единой… канонической модели». Такая модель «объясняет, почему молекулярно разнообразные нейронные системы демонстрируют одинаковое качественное поведение» (2015: 41) – и очевидно, является вопросом второго типа по Бэттерману. Ресурс, к которому обращается Чиримуута, - это «канонические нейронные вычисления» (КНВ):
вычислительные модули, которые применяют одни и те же фундаментальные операции в различных контекстах ... набор инструментов в виде вычислительных операций, которые мозг применяет к различным чувственным модальностям и анатомическим областям, и которые могут быть описаны на более высоких уровнях абстракции, нежели их биофизическая реализация. (Chirimuuta 2014: 138)
Примеры КНВ включают в себя ингибирование шунтирования, линейную фильтрацию, рекуррентное усиление и определение порогового значения. Вместо того, чтобы быть механизмами-набросками (которые ожидают поступления дополнительных механистических деталей, чтобы превратиться в полномасштабные механизмы "как оно на самом деле"), КНВ привлекают в другой объяснительный контекст, а именно – в тот, который задаётся вторым типом вопросов Бэттермана о причинах. Росс соглашается с каноническими моделями:
Чтобы понять подход динамических систем, который нейроучёные используют для объяснения поведения [системы], необходимо уделить внимание тому, что представляет для них интерес в качестве объясняемого, а также уникальным инструментам моделирования [например, каноническим моделям], распространенным в их области. (2015: 52)
Говоря вкратце, ответы Чиримууты и Росса на вызов, сформулированный Капланом и Крейвером, являются общими для философии: спасти конкретную форму объяснения от коллапсирования в другую форму, разделив то, что подлежит объяснению.
Наконец, завершая обсуждать возвышение механицизма, отметим, что в недавних дискуссиях вновь появился аналог проблемы Крейвера (Craver 2007) касательно учета «конститутивной механистической релевантности», - то есть определения, какие активные компоненты системы в действительности являются частью механизма для данного системного явления. Роберт Руперт (Rupert 2009) предполагает, что «интеграция» - это ключевой критерий для определения того, какой набор вносящих причинно-следственный вклад механизмов составляет систему для некоторой задачи (основываясь на относительной частоте, с которой множества механизмов совместно вносят вклад в возникновение задач). Он трактует эту частоту совместного вклада как вероятность, с которой множество [механизмов] приводит к возникновению когнитивной задачи, условной по отношению к любому другому возникающему причинно значимому множеству [механизмов]. Фелипе Де Бригар (De Brigard 2017) оспаривает критерий Руперта, утверждая, что тот не может объяснить когнитивные системы, демонстрирующие две особенности: «диахроническую динамичность» вместе с «функциональной стабильностью». Частота, с которой данный механизм вносит причинно-следственный вклад в одну и ту же когнитивную задачу (функциональная стабильность), может со временем меняться (диахроническая динамичность). Хотя посредством затейливого мысленного эксперимента Де Бригар подчеркивает принципиальную важность этих особенностей для критерия интеграции Руперта, он также утверждает, что оба они являются широко распространенным явлением в человеческом мозге. Обе эти особенности обнаруживаются, например, в данных, относящихся к «уменьшению асимметрии полушарий у пожилых людей»: в решении задач, которые задействуют расположенные на полушариях области префронтальной коры у молодых людей, заметно снижение асимметрии полушарий у пожилых людей. И то, и другое можно найти в работе «Задне-переднем смещении с возрастом (Posterior-Anterior Shift with Aging)», где предложенная задача увеличивает активность в передних отделах головного мозга при уменьшении активности в задних отделах у пожилых людей - по сравнению с активностью, вызываемой той же задачей у более молодых людей.
Чтобы заменить Рупертовское понятие интеграции (в качестве критерия, определяющего, какие наборы механизмов составляют когнитивную систему), Де Бригар указывает на два недавних многообещающих достижения в сетевой нейробиологии, которые потенциально позволяют параметризовывать время. «Масштабируемая инклюзивность» - это метод проверки каждого узла в сети и определения его принадлежности к «структурам сообщества» на разных итерациях данной сети. «Временно-динамический сетевой анализ» - это способ количественной оценки изменений в структурах или модулях сообщества между сетями в разные моменты времени. Таким образом, оба метода определяют «модульные альянсы», которые в одной модели передают информацию как о совместной активации, так и о динамических изменениях. Де Бригар предлагает рассмотреть именно эти методы в качестве претендентов на роль того, что позволяет идентифицировать когнитивные системы.
Ясно, что в отношении влияния [понятия] механизма, обретшего силы в философии нейронаук в течение последнего десятилетия, многое еще предстоит обсудить. Однако, хотя понятие механизма сформировало наиболее влиятельную общую перспективу в этой области, в других областях также продолжается работа. Майкл Андерсон защищает значение когнитивной нейробиологии для определения таксономии психологии, вне зависимости от какой бы то ни было приверженности механицизму. Наиболее подробно его подход развит в книге «После френологии» (Anderson 2014), основанной на его влиятельной гипотезе «повторного использования нейронов». Как было признано благодаря стандартным методам когнитивной нейробиологии (особенно фМРТ), каждая область мозга участвует в сильно различающихся между собой когнитивных функциях и образует разнообразные «нейронные партнерства» с другими частями мозга в разных обстоятельствах. Соответственно, тогда психологические категории должны быть переосмыслены в соответствии с выводами, предложенными в поддержку повторного использования нейронов на основе обширных эмпирических данных. Подлинная «постфренологическая» наука о сознании должна отбросить допущение, что каждый отдел мозга выполняет свои собственные фундаментальные вычисления. Тем самым работа Андерсона явно продолжает подпитывать постоянный интерес философии нейронаук к локализации когнитивных функций.
В более коротком обзоре Андерсон (Anderson 2015) исследует значение когнитивной нейробиологии для переосмысления основных категорий психологии, исходя из следствий его гипотезы о повторном использовании нейронов. Попытки отобразить когнитивные процессы на конкретные нейронные процессы и области мозга выявляют отношения «многие-ко-многим». В итоге эти отношения не только показывают, что комбинированные анатомо-функциональные метки для областей мозга (например, «веретенообразная область распознавания лица») являются обманчивыми; выявленные отношения также заставляют усомниться в возможности сделать выбор между альтернативными психологическими таксономиями, обращаясь к когнитивным нейронаучным данным.
По мнению всех, за исключением самых упрямых сторонников автономии психологии от нейробиологии, эти сопоставления «многие-ко-многим» предполагают, что привносимая нами в проект отображения (картографии функций) психологическая таксономия нуждается в пересмотре. Чтобы сделать такой вывод, не нужно быть ярым приверженцем эпистемологического превосходства когнитивной нейробиологии над психологией или пылать страстью к психоневральной редукции. Простого желания релевантности между когнитивной нейробиологией и категориями психологии вполне достаточно. Соответственно, эти дебаты касаются «требований к единой науке о сознании и подобающей роли нейробиологических доказательств в построении такой онтологии» (2015: 70), нежели легитимности той или иной дисциплины.
Андерсон делит ревизионистские проекты относительно психологии на три вида: он основывается на степени ревизионности, которую каждый из видов рекомендует для психологии, и диапазоне предлагаемых предсказанными ревизиями взаимно-однозначных сопоставлений функций-структур, которые станут доступны. «Консерваторы» полагают, что потребность в масштабном пересмотре базовой таксономии психологии будет сравнительно небольшой, - даже если принять во внимание больше нейронаучных данных, чем это делается в современных стандартных практиках. «Умеренные» настаивают на том, что наши знания о функциях мозга «могут (и должны) выступать в качестве единственного арбитра психологически реального» (2015: 70), главным образом путем «расщепления» или «объединения» используемых в настоящее время психологических концепций. «Радикалы» пророчат куда более значительные пересмотры даже самых простых понятий психологии – и даже после таких пересмотров они все еще не ожидают, что найдется много взаимно-однозначных отображений между областями мозга и новыми психологическими базовыми понятиями. Хотя Андерсон не подчеркивает эту связь (элиминативный материализм в течение двух десятилетий не казался многообещающим предприятием в философии сознания или нейронауках), читатели заметят сходство с темами, обсуждавшимися в Разделе 2 выше – только теперь целью радикальных ревизионистов становится научная, а не народная психология. Ключевым критерием того, что какая бы то ни было переформулировка в сфере когнитивной онтологии удовлетворительна, является степень поддержания ею выводов двух видов. «Прямые выводы» - от задействования определенной когнитивной функции до предсказания активности мозга; и «обратные выводы», - от наблюдения, что происходит активация конкретной области мозга или паттерна, до прогноза, что задействована конкретная когнитивная операция. В свете этого явного критерия Андерсон с пользой рассматривает работу ряда выдающихся психологов и когнитивных нейробиологов в каждой из выделенных им ревизионистских групп. Принимая во внимание более широкую приверженность Андерсона повторному использованию нейронов и предлагаемому им переходу на «вдохновленные эволюционизмом, экологические и энактивисткие термины»‹22›, собственные симпатии автора связаны с группой «Радикалов»
язык и математика, например, лучше всего будут поняты как расширение наших базовых способностей обработки аффордансов, дополняемых системами общественно используемых символов… Психологическая наука, которая возникает в результате проведённой переоценки, может сильно отличаться от той, которой мы занимаемся сегодня. (2015: 75)
Поворотные нейронаучные гипотезы продолжают быть в фокусе внимания современной философии нейронаук. Берит Брогаард (Brogaard 2012), к примеру, выступает за переосмысление стандартного «диссоцианистского» понимания Мелвином Гудейлом и Дэвидом Милнером (Goodale and Milner 1992) прославленных «двух потоков визуальной обработки» - поворотным, а ныне перекочевавшим в учебники результатом нейронаук конца ХХ века. Два компонента являются ключевыми в стандартном понимании диссоциации. Первый заключается в том, что информацию, релевантную для визуально управляемых моментальных действий (совершаемых «на лету») и для распознавания объектов, вычисляют различные области мозга, а именно – дорсальный тракт (который проходит от первичной зрительной коры через медиальную височную область к верхней и нижней теменной области долек) и вентральный тракт (который проходит от первичной зрительной коры через V4 и в нижнюю височную кору). Второй пункт состоит в том, что из этих двух составляющих только релевантная информация для распознавания визуальных объектов, обработанная в вентральном потоке, вносит вклад в характер осознанных визуальных переживаний.
Беспокойство Брогаард связано с тем, что приведённое выше стандартное понимание бросает вызов психофункционализму, на текущий момент –нашему наиболее правдоподобному «натуралистическому» подходу к ментальным состояниям. Подход психофункционализма относительно сознания опирается напрямую на нашу лучшую когнитивную психологию. Если φ - это некоторый тип ментального состояния, наследующий содержание визуального опыта, тогда, согласно когнитивной психологии, описанию φ удовлетворяет широкий спектр визуально-опосредованных убеждений и желаний, различные виды визуальных воспоминаний и т.д. Однако согласно тому же стандартному «диссоцианистскому» описанию двух визуальных потоков у Гудейла и Милнера, только состояния дорсального потока, а не состояния вентрального потока, представляют действительно эгоцентрические визуальные свойства, т.е. «реляционные свойства, объекты которых обретаются с точки зрения верующего или воспринимающего‹23›» (Brogaard 2012: 572). Но, согласно когнитивной психологии, состояния дорсального потока не играют столь обширной φ-роли. Тогда, согласно психофункционализму, «φ-ментальные состояния не способны представлять эгоцентрические свойства» (2012: 572). Но кажется «крайне правдоподобным», что некоторые из наших перцептивных убеждений и визуальных воспоминаний представляют эгоцентрические свойства. В таком случае перед нами дилемма: либо мы отвергаем психофункционализм (а вместе с ним и наш наиболее правдоподобный натуралистский проект по определению того, является ли конкретное ментальное состояние реализованным (instantiated)), либо мы отвергаем стандартную диссоцианистскую интерпретацию гипотезы Гудейла и Милнера о двух визуальных потоках, несмотря на обилие подтверждающих её эмпирических данных. Ни один из рогов приведённой дилеммы не выглядит приемлемым, хотя первый из них кажется лучше, поскольку психофункционализму как общей теории сознания не хватает той сильной эмпирической поддержки, какой располагает стандартная интерпретация гипотезы Гудейла и Милнера.
Тем не менее, Брогаард рекомендует сохранить психофункционализм и вместо этого отвергнуть «конкретную формулировку» гипотезы Гудейла и Милнера о двух визуальных потоках. Согласно интерпретации, которую предлагается отклонить, «информация из дорсального потока не может вносить вклад в потенциально сознательные репрезентации, вычисляемые вентральным потоком‹24›» (2012: 586–587) . Эгоцентрические репрезентации визуальной информации, вычисляемые дорсальным потоком, вносят вклад в осознаваемые репрезентации визуального потока «через соединения обратной связи» от нейронов дорсального до вентрального потоков (2012: 586). Это не отрицает диссоциации:
Информация об эгоцентрических свойствах объектов обрабатывается дорсальным потоком, а информация об аллоцентрических свойствах‹25› объектов обрабатывается вентральным потоком. (2012: 586)
Но эта гипотеза диссоциации «не имеет отношения к тому, какая информация передается частям мозга, которые обрабатывают информацию, коррелирующую с визуальным осознаванием» (2012: 586). С помощью этого переосмысления психофункционализм предстаёт согласуемым с двухпотоковой гипотезой Гудейла и Милнера (в которой дорсальному и вентральному потокам соответствуют типы информации о «Что» и «Где/Как») и множеством поддерживающих её эмпирических данных. Согласно Брогаард, психофункционализм в таком случае способен «правильно трактовать перцептивные и когнитивные состояния, несущие обработанную в вентральном зрительном потоке информацию, как способные представлять эгоцентрические свойства» (2012: 586).
Несмотря на то, что философия нейронаук продолжает фокусироваться на когнитивной / системной / вычислительной нейробиологии (см. соответствующее обсуждение в Разделе 7 выше), интерес к клеточному / молекулярному мейнстриму в нейробиологии, по-видимому, возрастает. Одна известная статья Энн-Софи Барвич и Карима Бшира (Barwich and Bschir 2017) представляет собой историко-философское исследование рецепторов, связанных с G-белком (GPCR). Работы о структуре и функциональной значимостью этих белков преобладали в молекулярной нейробиологии в течение последних сорока лет; их [белков] роль в механизмах различных когнитивных функций в настоящее время эмпирически задокументирована. И, тем не менее, в среде философов сложно обнаружить небольшой интерес или даже его проблеск относительно этого поворота в нейробиологическом мейнстриме. Добросовестные исторические исследования Барвич и Бшира об открытии и развитии этих научных объектов философски окупаются. Роль манипулируемости как критерия в пользу инструментального реализма в исследованиях науки в действии (science-in-practice) в лабораторных реалиях становится значимой «только после того, как ученые решили, как концептуально согласовать измеряемые следствия в точности с научным объектом» (2017: 1317). Научным объектам вроде GPCR приписывают различные степени реальности на разных этапах процесса открытия. Роль такого объекта в оценке реальности «соседних элементов исследования» также становится частью критериев реальности его самого.
Влияние науки в действии на философию науки в целом остро ощущалось в философии нейронаук: особенно это проявлялось в возросшем философском интересе к нейронаучным экспериментам. Само по себе это не должно удивлять; нейронаука в значительной степени опирается на лабораторные эксперименты, - особенно в рамках клеточного и молекулярного мейнстрима исследований, свойственного «Обществу нейронаук». Таким образом, призыв к пониманию эксперимента должен привлекать любого философа, решившегося на погружение в область клеточных / молекулярных основ нейронаук. Две статьи Жаклин Салливан (Sullivan 2009, 2010) оказались важны в акцентировании этой темы. В своей работе (Sullivan 2009) Салливан признает, что как Бикл (Bickle 2003), так и Крейвер (Craver 2007) фокусируются на клеточных и молекулярных механизмах долговременной потенциации, а также на зависящей от опыта форме синаптической пластичности. Однако, настаивает она, более обширные философские инвестиции, приводящие к «глобальным» подходам (Бикла - к безжалостному редукционизму, а Крейвера - к мозаичному единству), при этом затеняют важные аспекты реальной лабораторной нейронаучной практики. Она подчеркивает роль «субпротоколов», определяющих правильный способ сбора данных, в своей модели «экспериментального процесса» и иллюстрирует эти понятия на ряде примеров. Ее анализ показывает важное и недооцененное напряжение между парой общепринятых экспериментальных норм. Погоня за «надежностью» заставляет экспериментаторов глубже простраивать обширные лабораторные методы контроля. Погоня же за «внешней достоверностью» подталкивает их к обогащению экспериментальных сред, чтобы те становились более репрезентативно похожи на запутанную, «грязную» естественную среду за пределами лаборатории. Эти две нормы обычно противоречат друг другу: чтобы в большей степени следовать за одной, ученые вводят условия, которые ограничивают их в следовании за другой.
В другой своей статье (Sullivan 2010) Салливан предлагает подробную историю задачи водного лабиринта Морриса, прослеживая ее путь вплоть до оригинальных публикаций Морриса. Философы нейронаук безоговорочно предположили, что водный лвабиринт является широко распространенным нейронаучным протоколом в исследованиях пространственного обучения и памяти грызунов, однако подробная научная история не даёт столь ясной интерпретации. Научный комментарий относительно того, что измеряет эта задача (включая некоторые заметки самого Морриса) не раскрывается со временем и не приходит к четкому консенсусу. Салливан прослеживает источник этой научного неконсистености вплоть до влияния клеточно-молекулярного редукционизма эпохи 1980-х, побудившего возникновение в поведенческой нейробиологии экспериментальных протоколов, подобных водному лабиринту Морриса.
Нейробиолог Альсино Сильва, нейроинформатик Энтони Ландрет и философ нейронаук Джон Бикл (Silva, Landreth and Bickle 2014) сосредоточились на экспериментах, ведомые иной мотивацией. Все современные научные дисциплины растут головокружительными темпами; но, возможно, более всего это проявляется в нейронауках. Больше ни один ученый не в силах идти в ногу со всей соответствующей опубликованной литературой даже в своей узкой области исследований или полностью понять ее значение. Общее отсутствие ясности и согласия относительно того, что известно, что остается сомнительным и что было опровергнуто, создает специфические проблемы для планирования эксперимента. Существует признанная многими и неотложная необходимость в разработке стратегий и инструментов для решения этих проблем. Имея перед собой эту явную цель, книга Сильвы, Ландрета и Бикла описывает структуру и набор принципов того, как следует систематизировать опубликованные записи. Они выводят свои базовые схемы и принципы непосредственно из исследований поворотных случаев, имевших место в такой влиятельной нейронаучной области, как молекулярное и клеточное познание (MКП). Авторы описывают, как предлагаемая ими базовая схема может быть использована для создания карт экспериментальных результатов. Ученые, вооруженные этими картами исследований, затем смогут более эффективно определить, что было достигнуто в их областях, а где по-прежнему существуют пробелы в знаниях. Технология, необходимая для автоматической генерации этих карт, уже существует. Сильва, Ландрет и Бикл в общих чертах обрисовывают преобразующее, поистине революционное влияние, которое эти карты смогут оказать на современную науку.
Три цели мотивируют подход Сильвы, Ландрета и Бикла. Во-первых, они выводят свою базовую схему из клеточной и молекулярной нейробиологии обучения и памяти. Этот выбор был строго обусловлен знакомством с самой научной дисциплиной. Сильва сыграл важную роль в привнесении методов направленного воздействия на гены, применяемых к млекопитающим, в поведенческую нейробиологию; внимание же Бикла к безжалостно редуцирующей нейронауке зиждилось на этих и других экспериментальных результатах. И хотя каждый из различных видов экспериментов и принятых в их рамках доказательств был признан другими, цель авторов - стать первыми, кто систематизирует эту информацию, чтобы практикующим ученым легче было планировать эксперименты. Сильва, Ландрет и Бикл настаивают: важные новые эксперименты могут быть идентифицированы и спланированы, если методически заполнять различные формы доказательств, распознаваемые в их базовой схеме, а также применять различные формы экспериментов к пробелам в экспериментальной записи, выявленной в ходе этого процесса.
Во-вторых, Сильва, Ландрет и Бикл взяли прямой курс на проблему растущего количества, сложности и интеграции опубликованной литературы для планирования эксперимента. Они показывают, как графически взвешенные представления результатов исследований могут быть использованы как руководства для исследовательских решениями, и как их построить. Принципы построения этих карт - это принципы интеграции экспериментальных результатов, выведенные непосредственно из опубликованных исследований MКП. Используя конкретный пример из недавних достижений молекулярной нейробиологии, они показывают, как создавать небольшие карты, отражающие серию экспериментов, и как комбинировать эти маленькие карты, чтобы проиллюстрировать целую область исследований в области нейронауки.
Наконец, Сильва, Ландрет и Бикл начинают развивать научную дисциплину планирования эксперимента. Они предполагают, что причинно-следственные графики, составляющие их исследовательские карты, будут играть роль, аналогичную роли статистики в уже развитой науке анализа данных. Разработка такого ресурса может иметь серьезные последствия для дальнейшего развития индексов цитирования и других измерений влияния, позволяющих оценить вклад в научную область – от вкладов отдельных ученых до вкладов целых учреждений.
Совсем недавно Бикл и Костко (Bickle and Kostko 2018) расширили рамки схемы Сильвы, Ландрета и Бикла за пределы нейробиологии обучения и памяти. Их тематическое исследование основано на исследованиях в области развития и социальной нейробиологии, а именно - работах Майкла Мини и Моше Шайфа по эпигенетике поведения кормящих матерей грызунов в ответ на стрессовые реакции у потомства. Используя детали этого конкретного случая, Бикл и Костко развивают недоработанную в проекте Сильвы, Ландрета и Бикла идею, что эксперименты, специально сконструированные для своих полученных результатов, в случае успеха должны быть интегрированы непосредственно в уже существующий фон установленных результатов. И, утверждают авторы, такие эксперименты, «интегрируемые по замыслу» с другими, направлены не на установление доказательств индивидуальных причинно-следственных связей между нейробиологическими типами, а скорее – на формулирование целых причинных путей, связывающих множество явлений. Акцент авторов на причинных путях связан с подходом Лорен Росс (готовится к печати). Работа Росс особенно интересна в этом контексте, потому что она использует свою концепцию причинной связи в целях «причинно-следственной селекции», связанной с различием между фоновыми условиями и «истинными» (инициирующими) причинами некоторого представляющего интерес результата. Для Сильвы, Ландрета и Бикла (Silva, Landreth and Bickle 2014) учет этого различия также имеет решающее значение, и, чтобы обозначить это, они опираются на конкретный вид эксперимента с подключением (connection experiment) - «позитивные манипуляции». Обращение Бикла и Костко к причинно-следственным путям в детализированном конкретном исследовании, недавно проведённом в области нейробиологии развития, способно помочь связать более широкую работу Сильвы, Ландрета и Бикла по нейробиологическим экспериментам с работой Росс, основывающейся на биологии в целом.
Bibliography
(На русском)
• Кэндел Э. Р. Клеточные основы поведения / Пер. с англ. Н. Ю. Алексеенко, Г. И. Рожковой; Под ред. П. Г. Костюка, Д. А. Сахарова. М.: Мир, 1980.
• Нагель Т. Каково быть летучей мышью? / Пер. с англ. М.Эскиной в // Даглас Хофштадтер. Дэниел Деннетт. Глаз разума. Фантазии и размышления о самосознании и о душе. С.349-359. Самара: БАХРАХ-М, 2003.
• Сакс О. Человек, который принял жену за шляпу (и другие истории из врачебной практики). М.: АСТ, 2010
(На английском)
• Akins, Kathleen A., 1993a, “What Is It Like to Be Boring and Myopic?”, in Dennett and His Critics: Demystifying Mind, Bo Dahlbom (ed.), Cambridge, MA: Blackwell, 124–160.
• –––, 1993b, “A Bat without Qualities?”, in Readings in Mind and Language, Martin Davies and Glyn W. Humphreys (eds.), (Consciousness: Psychological and Philosophical Essays 2), Cambridge, MA: Blackwell Publishing, 258–273.
• –––, 1996, “Of Sensory Systems and the ‘Aboutness’ of Mental States:”, Journal of Philosophy, 93(7): 337–372. doi:10.2307/2941125
• Anderson, Michael L., 2014, After Phrenology: Neural Reuse and the Interactive Brain, Cambridge, MA: The MIT Press.
• –––, 2015, “Mining the Brain for a New Taxonomy of the Mind: A New Taxonomy of the Mind”, Philosophy Compass, 10(1): 68–77. doi:10.1111/phc3.12155
• Aston-Jones, Gary, Robert Desimone, J. Driver, Steven J. Luck, and Michael Posner, 1999, “Attention”, in Zigmond et al. 1999: 1385–1410.
• Balzer, Wolfgang, C. Ulises Moulines, and Joseph D. Sneed, 1987, An Architectonic for Science, Dordrecht: Springer Netherlands. doi:10.1007/978-94-009-3765-9
• Barwich, Ann-Sophie and Karim Bschir, 2017, “The Manipulability of What? The History of G-Protein Coupled Receptors”, Biology & Philosophy, 32(6): 1317–1339. doi:10.1007/s10539-017-9608-9
• Batterman, Robert W. and Collin C. Rice, 2014, “Minimal Model Explanations”, Philosophy of Science, 81(3): 349–376. doi:10.1086/676677
• Bechtel, William, 1998, “Representations and Cognitive Explanations: Assessing the Dynamicist’s Challenge in Cognitive Science”, Cognitive Science, 22(3): 295–318. doi:10.1207/s15516709cog2203_2
• Bechtel, William and Jennifer Mundale, 1999, “Multiple Realizability Revisited: Linking Cognitive and Neural States”, Philosophy of Science, 66(2): 175–207. doi:10.1086/392683
• Bechtel, William and Robert C. Richardson, 1993, Discovering Complexity: Decomposition and Localization as Strategies in Scientific Research, Princeton, NJ: Princeton University Press.
• Bechtel, William, Pete Mandik, Jennifer Mundale, and Robert Stufflebeam (eds.), 2001, Philosophy and the Neurosciences: A Reader, Malden, MA: Blackwell.
• Bermúdez, José Luis, 1998, The Paradox of Self-Consciousness, (Representation and Mind), Cambridge, MA: MIT Press.
• Bickle, John, 1992, “Revisionary Physicalism”, Biology & Philosophy, 7(4): 411–430. doi:10.1007/BF00130060
• –––, 1993, “Philosophy Neuralized: A Critical Notice of P.M. Churchland’s A Neurocomputational Perspective”, Behavior and Philosophy, 20(2): 75–88.
• –––, 1995, “Psychoneural Reduction of the Genuinely Cognitive: Some Accomplished Facts”, Philosophical Psychology, 8(3): 265–285. doi:10.1080/09515089508573158
• –––, 1998, Psychoneural Reduction: The New Wave, Cambridge, MA: MIT Press.
• –––, 2003, Philosophy and Neuroscience: A Ruthlessly Reductive Account, Norwell, MA: Springer Academic Publishers.
• ––– (ed.), 2009, The Oxford Handbook of Philosophy and Neuroscience, New York: Oxford University Press. doi:10.1093/oxfordhb/9780195304787.001.0001
• Bickle, John and Aaron Kostko, 2018, “Connection Experiments in Neurobiology”, Synthese, 195(12): 5271–5295. doi:10.1007/s11229-018-1838-0
• Biro, J. I., 1991, “Consciousness and Subjectivity”, in Consciousness, Enrique Villanueva (ed.), (Philosophical Issues 1), 113–133. doi:10.2307/1522926
• Bliss, T. V. P. and T. Lømo, 1973, “Long-Lasting Potentiation of Synaptic Transmission in the Dentate Area of the Anaesthetized Rabbit Following Stimulation of the Perforant Path”, The Journal of Physiology, 232(2): 331–356. doi:10.1113/jphysiol.1973.sp010273
• Block, Ned, 1987, “Advertisement for a Semantics for Psychology”, Midwest Studies in Philosophy, 10: 615–678. doi:10.1111/j.1475-4975.1987.tb00558.x
• –––, 1995, “On a Confusion about a Function of Consciousness”, Behavioral and Brain Sciences, 18(2): 227–247. doi:10.1017/S0140525X00038188
• Bogen, Jim, 2005, “Regularities and Causality; Generalizations and Causal Explanations”, Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, 36(2): 397–420. doi:10.1016/j.shpsc.2005.03.009
• Bower, James M. and David Beeman, 1995, The Book of GENESIS: Exploring Realistic Neural Models with the GEneral NEural SImulation System, New York: Springer-Verlag.
• Brogaard, Berit (Brit), 2012, “Vision for Action and the Contents of Perception”:, Journal of Philosophy, 109(10): 569–587. doi:10.5840/jphil20121091028
• Caplan, David N. , T. Carr, James L. Gould, and R. Martin, 1999, “Language and Communication”, in Zigmond et al. 1999: 1329–1352.
• Chalmers, David John, 1996, The Conscious Mind: In Search of a Fundamental Theory, (Philosophy of Mind Series), New York: Oxford University Press.
• Chirimuuta, M., 2014, “Minimal Models and Canonical Neural Computations: The Distinctness of Computational Explanation in Neuroscience”, Synthese, 191(2): 127–153. doi:10.1007/s11229-013-0369-y
• Churchland, Patricia Smith, 1986, Neurophilosophy: Toward a Unified Science of the Mind-Brain, (Computational Models of Cognition and Perception), Cambridge, MA: MIT Press.
• Churchland, Patricia Smith and Terrence J. Sejnowski, 1992, The Computational Brain, (Computational Neuroscience), Cambridge, MA: MIT Press.
• Churchland, Paul M., 1979, Scientific Realism and the Plasticity of Mind, Cambridge: Cambridge University Press. doi:10.1017/CBO9780511625435
• –––, 1981, “Eliminative Materialism and the Propositional Attitudes”, The Journal of Philosophy, 78(2): 67–90. doi:10.2307/2025900
• –––, 1987, Matter and Consciousness, revised edition, Cambridge, MA: MIT Press.
• –––, 1989, A Neurocomputational Perspective, Cambridge, MA: MIT Press.
• –––, 1995, The Engine of Reason, the Seat of the Soul, Cambridge, MA: MIT Press.
• –––, 1996, “The Rediscovery of Light”, The Journal of Philosophy, 93(5): 211–228. doi:10.2307/2940998
• Churchland, Paul M. and Patricia S. Churchland, 1997, “Recent Work on Consciousness: Philosophical, Theoretical, and Empirical”, Seminars in Neurology, 17(2): 179–186. doi:10.1055/s-2008-1040928
• Clark, Andy, 2016, Surfing Uncertainty: Prediction, Action, and the Embodied Mind, New York: Oxford University Press. doi:10.1093/acprof:oso/9780190217013.001.0001
• Clark, Austen, 1993, Sensory Qualities, Oxford: Oxford University Press. doi:10.1093/acprof:oso/9780198236801.001.0001
• Craver, Carl F., 2007, Explaining the Brain: What the Science of the Mind-Brain Could Be, Oxford: Oxford University Press. doi:10.1093/acprof:oso/9780199299317.001.0001
• Craver, Carl F. and Lindley Darden, 2001, “Discovering Mechanisms in Neurobiology”, in Machamer, Grush, and McLaughlin 2001: 112–137.
• Dennett, Daniel C., 1978, “Why You Can’t Make a Computer That Feels Pain”, Synthese, 38(3): 415–456. doi:10.1007/BF00486638
• –––, 1991, Consciousness Explained, New York: Little Brown.
• –––, 1995, “The Path Not Taken”, Behavioral and Brain Sciences, 18(2): 252–253. doi:10.1017/S0140525X00038243
• De Brigard, Felipe, 2017, “Cognitive Systems and the Changing Brain”, Philosophical Explorations, 20(2): 224–241. doi:10.1080/13869795.2017.1312503
• Dretske, Fred, 1981, Knowledge and the Flow of Information, Cambridge, MA: MIT Press.
• –––, 1988, Explaining Behavior, Cambridge, MA: MIT Press.
• Eliasmith, Chris, 2009, “Neurocomputational Models: Theory, Application, Philosophical Consequences”, in Bickle 2009: 346–369. doi:10.1093/oxfordhb/9780195304787.003.0014
• Eliasmith, Chris and Charles H. Anderson, 2003, Neural Engineering: Computation, Representation, and Dynamics in Neurobiological Systems, (Computational Neuroscience), Cambridge, MA: MIT Press.
• Farah, Martha J., 2008, “Neuroethics and the Problem of Other Minds: Implications of Neuroscience for the Moral Status of Brain-Damaged Patients and Nonhuman Animals”, Neuroethics, 1(1): 9–18. doi:10.1007/s12152-008-9006-8
• Farah, Martha J. and Paul Root Wolpe, 2004, “Monitoring and Manipulating Brain Function: New Neuroscience Technologies and Their Ethical Implications”, The Hastings Center Report, 34(3): 35–45.
• Feyerabend, Paul K., 1963, “Comment: Mental Events and the Brain”, The Journal of Philosophy, 60(11): 295–296. doi:10.2307/2023030
• Flanagan, Owen, 2009, “Neuro‐Eudaimonics or Buddhists Lead Neuroscientists to the Seat of Happiness”, in Bickle 2009: 582–600. doi:10.1093/oxfordhb/9780195304787.003.0024
• Fodor, Jerry A., 1974, “Special Sciences (or: The Disunity of Science as a Working Hypothesis)”, Synthese, 28(2): 97–115. doi:10.1007/BF00485230
• –––, 1981, RePresentations, Cambridge, MA: MIT Press.
• –––, 1987, Psychosemantics, Cambridge, MA: MIT Press.
• Fodor, Jerry and Ernest LePore, 1992, Holism: A Shopper’s Guide, Cambridge, MA: MIT Press.
• Gazzaniga, Michael S. (ed.), 1995, The Cognitive Neurosciences, Cambridge, MA: MIT Press.
• Friston, Karl and Stefan Kiebel, 2009, “Predictive Coding under the Free-Energy Principle”, Philosophical Transactions of the Royal Society B: Biological Sciences, 364(1521): 1211–1221. doi:10.1098/rstb.2008.0300
• Georgopoulos, A. P., A. B. Schwartz, and R. E. Kettner, 1986, “Neuronal Population Coding of Movement Direction”, Science, 233(4771): 1416–1419. doi:10.1126/science.3749885
• Goldman, Alvin I., 2006, Simulating Minds: The Philosophy, Psychology, and Neuroscience of Mindreading, New York: Oxford University Press. doi:10.1093/0195138929.001.0001
• Goodale, Melvyn A. and A. David Milner, 1992, “Separate Visual Pathways for Perception and Action”, Trends in Neurosciences, 15(1): 20–25.
• Grush, Rich, 2001, “The Semantic Challenge to Computational Neuroscience”, in Machamer, Grush, and McLaughlin 2001: 155–172.
• –––, 2004, “The Emulation Theory of Representation: Motor Control, Imagery, and Perception”, Behavioral and Brain Sciences, 27(3): 377–396. doi:10.1017/S0140525X04000093
• –––, 2005, “Brain Time and Phenomenological Time”, in Cognition and the Brain: The Philosophy and Neuroscience Movement, Andrew Brook and Kathleen Akins (eds.), Cambridge: Cambridge University Press, 160–207. doi:10.1017/CBO9780511610608.006
• Haken, H., J. A. S. Kelso, and H. Bunz, 1985, “A Theoretical Model of Phase Transitions in Human Hand Movements”, Biological Cybernetics, 51(5): 347–356. doi:10.1007/BF00336922
• Hardcastle, Valerie Gray, 1997, “When a Pain Is Not”, The Journal of Philosophy, 94(8): 381–409. doi:10.2307/2564606
• Hardin, C.L., 1988, Color for Philosophers: Unweaving the Rainbow, Indianapolis, IN: Hackett.
• Haugeland, John, 1985, Artificial Intelligence: The Very Idea, Cambridge, MA: MIT Press.
• Hawkins, Robert D. and Eric R. Kandel, 1984, “Is There a Cell-Biological Alphabet for Learning?” Psychological Review, 91(3): 375–391.
• Hebb, D.O., 1949, The Organization of Behavior: A Neuropsychological Theory, New York: Wiley.
• Hirstein, William, 2005, Brain Fiction: Self-Deception and the Riddle of Confabulation, Cambridge, MA: MIT Press.
• Hodgkin, Alan L., Andrew F. Huxley, and Bernard Katz, 1952, “Measurement of Current‐voltage Relations in the Membrane of the Giant Axon of Loligo”, The Journal of Physiology, 116(4): 424–448. doi:10.1113/jphysiol.1952.sp004716
• Hohwy, Jakob, 2013, The Predictive Mind, New York: Oxford University Press. doi:10.1093/acprof:oso/9780199682737.001.0001
• Hooker, C.A., 1981a, “Towards a General Theory of Reduction. Part I: Historical and Scientific Setting”, Dialogue, 20(1): 38–59. doi:10.1017/S0012217300023088
• –––, 1981b, “Towards a General Theory of Reduction. Part II: Identity in Reduction”, Dialogue, 20(2): 201–236. doi:10.1017/S0012217300023301
• –––, 1981c, “Towards a General Theory of Reduction. Part III: Cross-Categorical Reduction”, Dialogue, 20(3): 496–529. doi:10.1017/S0012217300023593
• Horgan, Terence and George Graham, 1991, “In Defense of Southern Fundamentalism”, Philosophical Studies, 62(2): 107–134. doi:10.1007/BF00419048
• Hubel, D. H. and T. N. Wiesel, 1962, “Receptive Fields, Binocular Interaction and Functional Architecture in the Cat’s Visual Cortex”, The Journal of Physiology, 160(1): 106–154. doi:10.1113/jphysiol.1962.sp006837
• Jackson, Frank and Philip Pettit, 1990, “In Defence of Folk Psychology”, Philosophical Studies, 59(1): 31–54. doi:10.1007/BF00368390
• Kaplan, David Michael and Carl F. Craver, 2011, “The Explanatory Force of Dynamical and Mathematical Models in Neuroscience: A Mechanistic Perspective*”, Philosophy of Science, 78(4): 601–627. doi:10.1086/661755
• Kawato, M., 1999, “Internal Models for Motor Control and Trajectory Planning”, Current Opinion in Neurobiology, 9(6): 718–727.
• Klein, C., 2010, “Images Are Not the Evidence in Neuroimaging”, The British Journal for the Philosophy of Science, 61(2): 265–278. doi:10.1093/bjps/axp035
• Kolb, Bryan and Ian Q. Whishaw, 1996, Fundamentals of Human Neuropsychology, 4th edition, New York: W.H. Freeman.
• Kosslyn, Stephen M., 1997, “Mental Imagery”, in Conversations in the Cognitive Neurosciences, Michael S. Gazzaniga (ed.), Cambridge, MA: MIT Press, pp. 37–52.
• Lee, Choongkil, William H. Rohrer, and David L. Sparks, 1988, “Population Coding of Saccadic Eye Movements by Neurons in the Superior Colliculus”, Nature, 332(6162): 357–360. doi:10.1038/332357a0
• Lehky, Sidney R. and Terrence J. Sejnowski, 1988, “Network Model of Shape-from-Shading: Neural Function Arises from Both Receptive and Projective Fields”, Nature, 333(6172): 452–454. doi:10.1038/333452a0
• Lettvin, J., H. Maturana, W. McCulloch, and W. Pitts, 1959, “What the Frog’s Eye Tells the Frog’s Brain”, Proceedings of the IRE, 47(11): 1940–1951. doi:10.1109/JRPROC.1959.287207
• Levine, Joseph, 1983, “Materialism and Qualia: The Explanatory Gap”, Pacific Philosophical Quarterly, 64(4): 354–361. doi:10.1111/j.1468-0114.1983.tb00207.x
• Levy, Neil, 2007, Neuroethics: Challenges for the 21st Century, Cambridge: Cambridge University Press. doi:10.1017/CBO9780511811890
• Llinás, Rodolfo R., 1975, “The Cortex of the Cerebellum”, Scientific American, 232(1/January): 56–71. doi:10.1038/scientificamerican0175-56
• Llinás, Rodolfo R. and Patricia Smith Churchland (eds.), 1996, The Mind-Brain Continuum: Sensory Processes, Cambridge, MA: MIT Press.
• Lloyd, Dan, 2002, “Functional MRI and the Study of Human Consciousness”, Journal of Cognitive Neuroscience, 14(6): 818–831. doi:10.1162/089892902760191027
• –––, 2003, Radiant Cool: A Novel Theory of Consciousness, Cambridge, MA: MIT Press.
• Machamer, Peter, Lindley Darden, and Carl F. Craver, 2000, “Thinking about Mechanisms”, Philosophy of Science, 67(1): 1–25. doi:10.1086/392759
• Machamer, Peter K., Rick Grush, and Peter McLaughlin (eds.), 2001, Theory and Method in the Neurosciences (Pittsburgh-Konstanz Series in the Philosophy and History of Science), Pittsburgh, PA: University of Pittsburgh Press.
• Magistretti, Pierre J., 1999, “Brain Energy Metabolism”, in Zigmond et al. 1999: 389–413.
• Nagel, Thomas, 1971, “Brain Bisection and the Unity of Consciousness”, Synthese, 22(3–4): 396–413. doi:10.1007/BF00413435
• –––, 1974, “What Is It Like to Be a Bat?”, The Philosophical Review, 83(4): 435–450. doi:10.2307/2183914
• Piccinini, Gualtiero and Carl Craver, 2011, “Integrating Psychology and Neuroscience: Functional Analyses as Mechanism Sketches”, Synthese, 183(3): 283–311. doi:10.1007/s11229-011-9898-4
• Place, U. T., 1956, “Is Consciousness a Brain Process?”, British Journal of Psychology, 47(1): 44–50. doi:10.1111/j.2044-8295.1956.tb00560.x
• Polger, Thomas W., 2004, Natural Minds, Cambridge, MA: MIT Press.
• Prinz, Jesse J., 2004, Gut Reactions: A Perceptual Theory of the Emotions. New York: Oxford University Press.
• –––, 2007, The Emotional Construction of Morals, Oxford: Oxford University Press. doi:10.1093/acprof:oso/9780199571543.001.0001
• Putnam, Hilary, 1967, “Psychological Predicates”, in Art, Mind, and Religion: Proceedings of the 1965 Oberlin Colloquium in Philosophy, W. H. Capitan and D. D. Merrill (eds.), Pittsburgh, PA: University of Pittsburgh Press, pp. 49–54.
• Rall, Wilfrid, 1959, “Branching Dendritic Trees and Motoneuron Membrane Resistivity”, Experimental Neurology, 1(5): 491–527. doi:10.1016/0014-4886(59)90046-9
• Ramsey, William, 1992, “Prototypes and Conceptual Analysis”, Topoi, 11(1): 59–70. doi:10.1007/BF00768299
• Roskies, Adina L., 2007, “Are Neuroimages Like Photographs of the Brain?”, Philosophy of Science, 74(5): 860–872. doi:10.1086/525627
• –––, 2009, “What’s ‘Neu’ in Neuroethics?”, in Bickle 2009: 454–472. doi:10.1093/oxfordhb/9780195304787.003.0019
• Ross, Lauren N., 2015, “Dynamical Models and Explanation in Neuroscience”, Philosophy of Science, 82(1): 32–54. doi:10.1086/679038
• –––, forthcoming, “Causal Concepts in Biology: How Pathways Differ from Mechanisms and Why It Matters”, The British Journal for the Philosophy of Science, first online: 12 December 2018. doi:10.1093/bjps/axy078
• Rumelhart, D. E., G. E. Hinton, and J. L. McClelland, 1986, “A Framework for Parallel Distributed Processing”, in Parallel Distributed Processing: Explorations in the Microstructure of Cognition, volume 1, D. E. Rumelhart and J. L. McClelland (eds.), Cambridge, MA: MIT Press, pp. 45–76.
• Rupert, Robert D., 2009, Cognitive Systems and the Extended Mind, New York: Oxford University Press. doi:10.1093/acprof:oso/9780195379457.001.0001
• Schaffner, Kenneth F., 1992, “Philosophy of Medicine”, in Introduction to the Philosophy of Science: A Text by Members of the Department of the History and Philosophy of Science of the University of Pittsburg, Merrilee H. Salmon, John Earman, Clark Glymour, James G. Lennox, Peter Machamer, J. E. McGuire. John D. Norton, Wesley C. Salmon, and Kenneth F. Schaffner (eds.), Englewood Cliffs, NJ: Prentice-Hall, pp. 310–345.
• Schechter, Elizabeth, 2018, Self-Consciousness and ‘Split’ Brains: The Minds’ I, New York: Oxford University Press. doi:10.1093/oso/9780198809654.001.0001
• Schneider, Susan, 2009, “Future Minds: Transhumanism, Cognitive Enhancement, and the Nature of Persons”, in Penn Center Guide to Bioethics, Vardit Ravitsky, Autumn Fiester, and Arthur L. Caplan (eds.), New York: Springer, pp. 95–110.
• Schroeder, Timothy, 2004, Three Faces of Desire, Oxford: Oxford University Press. doi:10.1093/acprof:oso/9780195172379.001.0001
• Silberstein, Michael and Anthony Chemero, 2013, “Constraints on Localization and Decomposition as Explanatory Strategies in the Biological Sciences”, Philosophy of Science, 80(5): 958–970. doi:10.1086/674533
• Silva, Alcino J., Anthony Landreth, and John Bickle, 2014, Engineering the Next Revolution in Neuroscience: The New Science of Experiment Planning, New York: Oxford University Press. doi:10.1093/acprof:oso/9780199731756.001.0001
• Smart, J. J. C., 1959, “Sensations and Brain Processes”, The Philosophical Review, 68(2): 141–156. doi:10.2307/2182164
• Stich, Stephen, 1983, From Folk Psychology to Cognitive Science, Cambridge, MA: MIT Press.
• Stufflebeam, Robert S. and William Bechtel, 1997, “PET: Exploring the Myth and the Method”, Philosophy of Science, 64(supplement/December): S95–S106. doi:10.1086/392590
• Sullivan, Jacqueline A., 2009, “The Multiplicity of Experimental Protocols: A Challenge to Reductionist and Non-Reductionist Models of the Unity of Neuroscience”, Synthese, 167(3): 511–539. doi:10.1007/s11229-008-9389-4
• –––, 2010, “Reconsidering ‘Spatial Memory’ and the Morris Water Maze”, Synthese, 177(2): 261–283. doi:10.1007/s11229-010-9849-5
• Suppe, Frederick, 1974, The Structure of Scientific Theories, Urbana, IL: University of Illinois Press.
• Tye, Michael, 1993, “Blindsight, the Absent Qualia Hypothesis, and the Mystery of Consciousness”, in Philosophy and the Cognitive Sciences, Christopher Hookway and Donald M. Peterson (eds.), (Royal Institute of Philosophy Supplement 34), Cambridge: Cambridge University Press, 19–40. doi:10.1017/S1358246100002447
• Van Fraassen, Bas C., 1980, The Scientific Image, New York: Oxford University Press. doi:10.1093/0198244274.001.0001
• Von Eckardt Klein, Barbara, 1975, “Some Consequences of Knowing Everything (Essential) There Is to Know About One’s Mental States”, The Review of Metaphysics, 29(1): 3–18.
• –––, 1978, “Inferring Functional Localization from Neurological Evidence”, in Explorations in the Biology of Language, Edward Walker (ed.), Cambridge, MA: MIT Press, pp. 27–66.
• Weber, Marcel, 2008, “Causes without Mechanisms: Experimental Regularities, Physical Laws, and Neuroscientific Explanation”, Philosophy of Science, 75(5): 995–1007. doi:10.1086/594541
• Woodward, James, 2003, Making Things Happen: A Theory of Causal Explanation, Oxford: Oxford University Press. doi:10.1093/0195155270.001.0001
• Zigmond, Michael J., Floyd E. Bloom, Story C. Landis, James L. Roberts, and Larry R. Squire (eds.), 1999, Fundamental Neuroscience, San Diego, CA: Academic Press.









